A study of feedback loop mechanisms regulating calcium, IP3 and dopamine in neurons
Anand Pawar
, ∗
,
Kamal Raj Pardasani
Expand
Department of Mathematics, Bioinformatics and Computer Applications, Maulana Azad National Institute of Technology, Bhopal-462003, Madhya Pradesh, India
Author to whom any correspondence should be addressed.
Anand Pawar, Kamal Raj Pardasani. A study of feedback loop mechanisms regulating calcium, IP3 and dopamine in neurons[J]. Communications in Theoretical Physics, 2025, 77(6): 065001. DOI: 10.1088/1572-9494/ada495
Introduction
An assortment of cellular processes, including but not limited to diffusion, buffering, influx, and outflux mechanisms, play a role in the complex and versatile bioenergetic systems that facilitate chemical signaling. This process is observed in various cells like neurons and is mediated by calcium, IP3, and dopamine signaling molecules. Neuronal cell calcium signaling is essential for the regulation of numerous cellular processes. Neuronal disorders may manifest due to any disruption in the calcium signaling pathway. Alzheimer's disease (AD) has been associated with alterations in calcium buffering [1], hyperregulation of the sarcoplasmic/endoplasmic reticulum calcium ATPase (SERCA) pump and leak activities [2], and heightened sensitivity of calcium release channels [3]. Nevertheless, under realistic circumstances, calcium regulation depends on the interplay of additional signaling systems, including IP3 and DA, which occur within neuronal cells. The IP3 molecule facilitates the increase of cytosolic Ca2+ levels by discharging Ca2+ ions from the endoplasmic reticulum (ER) to the cytosol; it operates as a secondary messenger. Multiple cellular processes, including cell proliferation, fertilization, cell transformation, and muscular contraction, are intricately regulated by this process. Dopamine is a neurochemical messenger that has a significant concentration inside the midbrain region of the central nervous system [4]. Dopamine is a potentially harmful chemical that has been associated with the onset of neurodegenerative conditions like Parkinson's disease (PD) [5]. Dopamine is influenced by a wide range of metabolites, many biological constituents, and a diverse array of environmental and genetic influences in neuronal cells. The mathematical investigation of DA signaling and its potential relevance to Parkinson's illness has been reported in cells [6].
The cell utilizes two separate sources of calcium ions to generate signals: the discharge of Ca2+ ions from internal cellular compartments and the Ca2+ inflow across the plasma membrane. Both the processes of releasing and entering Ca2+ can produce highly localized Ca2+ signals, which are regarded as essential events in Ca2+ signaling [7]. The ryanodine receptor (RyR) and IP3-receptor (IP3R) are distinct Ca2+-releasing channels positioned within the ER membrane [8]. The calcium buffering mechanism involves the use of both stationary and mobile buffers, which significantly affects the movement of calcium ions in cellular settings [9]. A calcium buffer was used to induce a quantifiable decrease in the alteration of free calcium levels following a transient infusion of calcium [10]. Recent studies examined the phenomenon of calcium signaling and its modulation in many types of human cells, such as neurons [11–16], astrocytes [17, 18], acinar [19, 20], myocyte [21, 22], Oocyte [23–27], fibroblast cells [28, 29], hepatocyte [30–32] and T-lymphocyte [33]. An investigation has been carried out on the process of diffusion of calcium ions within neuron cells using the excess buffer approximation as a theoretical framework. This framework includes the impact of sodium (Na+) ions on the movement of calcium, while also including different sources of Ca2+ and Na+ in neurons [34]. The study investigated calcium diffusion using a two-dimensional model, taking into consideration several elements such as buffer, diffusion coefficients, and flux, to accurately assess their impacts [35]. The external environment has a crucial impact on the regulation of cytosolic Ca2+ levels in neuronal cells by governing the inflow and outflow of Ca2+ ions through the plasma membrane [36].
There has been a lack of extensive research on the connections between calcium and IP3 systems in various types of human cells, both in experimental and theoretical settings. The findings obtained by analyzing interconnected signaling systems such as Ca2+ and IP3 reveal significant disparities when compared to those obtained from examining separate signaling systems such as Ca2+ in neuronal cells. The structure and functioning of IP3-mediated Ca2+ signaling have a substantial impact on the control of brain activity [37]. Observations have been made about the link between fluctuations and chaotic dynamics in Ca2+ signaling, as well as the coexistence of dynamic calcium and constant IP3 within neural cells [38]. A study has reported a nine-variable kinetic model that accurately describes the behavior of the IP3R channel by using quantitative measurements of Ca2+ ions activation and inhibition of the IP3-receptor situated in the ER [39]. The nine-variable IP3R model has been analyzed and simplified, resulting in a two-variable system [40]. Measuring the transportation coefficient of IP3 and Ca2+ is important for determining the extent of messenger action from a localized source of calcium and IP3 inside cellular contexts [41]. The interplay between the concentration of calcium ions and inositol trisphosphate results in the creation of a state of bistability. This is achieved by a feedback process where Ca2+ concentration influences the formation of IP3, and the discharge of calcium ions is regulated by both calcium ions and IP3 [42]. The investigations undertaken on the calcium and IP3 signaling systems have been the subject of recent reports. These investigations have revealed their role in regulating many mechanisms in different cell types, such as neurons [43, 44], hepatocytes [45], β-cell [46], fibroblast cells [47, 48], T-lymphocyte [49], and others. Prior studies have examined the exploration of the interrelated spatial and temporal impacts of Ca2+ and IP3 mechanics, together with their contributions to the generation of nitric oxide [50] and β-amyloid [51], and the release of ATP [52] in neuronal cells.
Several calcium-activated channels, including voltage-gated Ca2+ channels (VGCC) [53], are involved in controlling the distinct functions of dopamine neuron cells. PD is identified by the deterioration of dopaminergic neurons, which is influenced by high levels of calcium, dopamine, and α-synuclein [54]. The function of the Calbindin-D28K buffer in safeguarding dopaminergic neuronal cells against the pathological process that causes PD was elucidated in the literature [55]. The increase in Ca2+ ion concentration can be attributed to the presence of a small quantity of calbindin-D28k, which is both indicative and characteristic of Parkinson's disease [56]. Several mechanisms such as source influx, buffering, ryanodine receptor activity, and other factors have been identified as contributors to the high levels of calcium and dopamine that eventually result in the death of dopaminergic neurons [57].
Utilizing fractional calculus in the analysis of biological systems provides a more profound understanding of the intricate mechanisms exhibited by different cells and tissues [58]. Various numerical approaches such as the Grunwald approximation are employed to solve the fractional derivatives along spatial dimensions [59]. The fractional model is utilized to elucidate memory phenomena across several disciplines, encompassing mechanics, biology, and psychology [60]. Cell memory is commonly linked to fractional-order systems, and cells acquire learning through their encounters with disease-related circumstances [61]. A notable finding reported in the literature is that the memory trace begins to emerge when the fractional order increases from 0 to 1 and entirely vanishes when the order reaches 1 [62]. The Ca2+ dynamics within neurons are characterized using a fractional reaction-diffusion framework that incorporates the presence of calcium-binding proteins [63]. Cell memory is found to have a notable influence on protein and VGCC. Cellular memory reduces the influx of calcium ions with elevation, leading to improved outcomes in estimating the Ca2+ flow during pathological conditions [64]. Superdiffusion is a rare occurrence that can be observed in several situations, such as biological systems and the motion of organic molecules [65]. The augmentation of the diffusion mechanism is noted when the fractional derivatives are employed as replacements for the second-order derivatives inside the diffusion framework. The phenomenon characterized by an enhanced rate of diffusion is commonly referred to as superdiffusion [66]. The studies on the effects of memory and superdiffusion on the interacting signaling of Ca2+ and IP3 [67], Ca2+ and β-amyloid [68], Ca2+ and NO [69], Ca2+ and DA [70], and Ca2+, IP3, and β-amyloid [71] in neurons have been analyzed with numerical findings. These investigations have revealed a significant discrepancy in results when comparing systems with fractional-order dynamics to those with integer-order dynamics.
The existing integer-order models of Ca2+, IP3, and DA systems in the literature exhibit mutual interactions among these signaling systems in neuronal cells, providing significant insights into many biological functions within neuronal cells. However, there is no documented research on the interaction among Ca2+, IP3, and DA in nonlinear fractional-order dynamics, which illustrates the impacts of superdiffusion and cellular memory with Brownian motion (BM) in neuronal cells. The principal advantage of examining the space-time fractional reaction-diffusion equation compared to the classical diffusion equation is its global behavior. The integer-order differential equation acts as a local operator, while the fractional (non-integer) order differential equation performs as a non-local operator. Estimating the spatial and temporal profiles of Ca2+, IP3, and DA in neurons in any certain state depends not only on the current profiles of these molecules but also on all prior profiles of Ca2+, IP3, and DA. The dynamics of the fractional reaction-diffusion equation facilitate the calculation of precise concentration profiles for Ca2+, IP3, and DA clarifying the complex mechanisms involved in the nervous system. Therefore, a fractional-order reaction-diffusion model of interacting Ca2+, IP3, and DA has been developed in neurons. This paper presents the framework of a feedback loop model with fractional order to accurately represent neuronal calcium, IP3, and DA signaling, which interact with nonlinear dynamics. The computational method utilized in this study involves the implementation of the Crank–Nicholson (CN) scheme in conjunction with the Grunwald technique for spatial dimensions. Additionally, the L1 scheme is applied for temporal dimensions, and Gauss–Seidel Iterations (GSI) is used for the solution process. The feedback loop framework incorporates the reciprocal relationship between Ca2+ and IP3, the one-way relationship between Ca2+ and DA, and the one-way relationship between DA and IP3, along with various crucial such as VGCC, plasma membrane Ca2+ ATPase (PMCA) channel, sodium–calcium exchanger (NCX), buffer, STIM-Orai channel, RyR, IP3R, etc in neuronal cells. This study investigates the impact of various significant processes such as superdiffusion and memory characterized by BM on the interacting calcium, IP3, and DA signaling in neuronal cells providing novel insights into these impacts.
Mathematical formulation
The existing integer-order Ca2+ model proposed by Wagner et al [42] has been extended to fractional order along both spatial and temporal dimensions. This extended model also incorporates the EGTA buffer, VGCC, STIM-Orai, PMCA, NCX, and RyR into the fractional-order Ca2+ framework and can be depicted as follows:
In this context, $0\lt {U}_{1}\leqslant 1,1\lt {V}_{1}\leqslant 2.$ The parameters [B]∞ and [Ca2+]∞ denote the equilibrium concentrations of buffer and Ca2+ ions in neuronal cells, respectively. The variables t and x are employed to denote the temporal and spatial dimensions, respectively. The diffusion coefficient of calcium ions is denoted as DCa, while the rate at which calcium ions associate with buffers is represented by K+.
The flux of IP3-receptor (JIPR) denotes the process by which calcium is released from ER to the cytosol of neuronal cells in response to IP3 stimulation as follows [42],
The parameters KIP3, KAc, and Kinh stand for the dissociation constants associated with the binding location of inositol trisphosphate and the activation and inhibition of Ca2+ in neuronal cells.
The SERCA pump flux (JSERCA) depicts the SERCA pump activity, which causes the translocation of cytosolic calcium ions from neuronal cells to the ER. The mathematical representation for the JSERCA is as follows [42],
In this context, the variables VSERCA and KSERCA are employed to denote the rate at which flux occurs and the constant that characterizes the Michaelis–Menten kinetics, respectively, for the SERCA pump.
The leak flux (JLeak) exhibits the leakage event, in which the gradual diffusion of Ca2+ ions from the ER membrane to the cytoplasm, occurs in the following manner [42],
In this context, the variable VRyR is employed to denote the flux rate constant associated with the RyR, while P0 represents the open probability of the RyR that ranges from 0 to 1.
The current equation proposed by Goldmann–Hodgkin–Kartz is utilized to simulate the flow of voltage-gated calcium channels (JVGCC) as follows,
Here, equation (8) incorporates the extracellular concentration of calcium ions ([Ca2+]0), the intracellular concentration of calcium ions ([Ca2+]i), the permeability of calcium ions (PCa), valency of calcium ions (ZCa), membrane potential (Vm), Faraday's parameter (F), real gas constant (R), and absolute temperature (T). The mathematical representation for converting equation (8) into the unit of molar/second is depicted in the following manner,
In neural cells, The NCX enables the transfer of a single calcium ion in exchange for three Na+ ions. The following is the mathematical representation for the NCX,
where, the variables Nai and Na0 represent the respective levels of intracellular and extracellular Na+ ions.
The PMCA pump flux (JPMCA) exhibits the activities of the PMCA pump, which facilitates the removal of cytosolic calcium from the intracellular environment to the extracellular space in the following manner,
In this context, the parameters VPMCA and KPMCA are employed to depict the flux rate constant and the dissociation pump rate for the PMCA, respectively.
The Orai channel (JOrai) flux exhibits the Orai channel activities, which facilitates the transfer of Ca2+ from the external environment to the cytosol in the following manner,
The existing integer-order IP3 framework proposed by Wagner et al [42] has been extended to fractional order along both spatial and temporal dimensions. This extended model also incorporates the Ca2+-dependent and DA-dependent flux of IP3 production into the fractional-order IP3 model and can be expressed as follows:
where $0\lt {U}_{2}\leqslant 1,1\lt {V}_{2}\leqslant 2.$ The IP3 diffusion coefficient is exhibited by Di. The term Jproduction is used to symbolize the flux of calcium-dependent inositol trisphosphate generation in as follows [42],
Here, the IP3 generation rate and the Michaelis parameter concerning the activation of Ca2+ are denoted as VProduction and KProduction, respectively. The mathematical expressions for the flux of degradation of inositol trisphosphate by JKinase [72] and Jphosphatase [73] are depicted as follows,
Here, the parameters associated with low and high concentrations of calcium ions (3-kinase) are denoted as Va and Vb, respectively. Additionally, Vph represents the maximal rate constant of the phosphatase. The flexible parameter λ depicts the elimination rate of IP3 [42].
The process of IP3 synthesis through the activation of D1-receptor and its reliance on dopamine can be represented by the following mathematical expressions,
where the maximum formation rate of IP3 by phospholipase C (PLC) due to D1-receptor activation is denoted as VDA. The binding strength of DA to the receptor is represented by the affinity constant, KDA.
The existing integer-order DA model proposed by Tello-Bravo [74] has been extended to fractional order along both spatial and temporal dimensions. This extended model also incorporates the Ca2+-dependent DA production into the fractional-order DA model and can be expressed as follows:
Here, $0\lt {U}_{3}\leqslant 1,1\lt {V}_{3}\leqslant 2.$ The parameter DDA represents the DA diffusion coefficient. Equation (20) incorporates the dopamine release per electrical stimulus pulse (DAP), stimulus frequency (f), Michaelis parameter (Km) and DA uptake rate (Vmax). The formulation of [Ca2+]-dependent dopamine release is presented as follows [74],
where the term Jrelease represents the flux of DA release. Equation (21) incorporates the average release flux ($\Psi$), high release probability (Prel) and Ca2+ sensitivity (Krel) in neuronal cells.
Initial conditions
The initial concentration for the Ca2+ [75], inositol trisphosphate [76], and dopamine [77] are shown as follows,
Equations (1)–(30) present the initial boundary value problem (IBVP) formulation for fractional-order PDEs, governing the interacting systems of Ca2+, IP3 and dopamine. The numerical methodology and analysis of stability have been elucidated in the appendix.
Results and discussion
The graphical representation of the numerical results pertaining to the feedback loop mechanisms of Ca2+, IP3, and dopamine systems with fractional-order dynamics across spatiotemporal dimensions has been presented. This graphical depiction aids in comprehending various essential cellular processes such as superdiffusion and memory displaying Brownian motion within neuronal cells. Table 1 displays a list of several parameters alongside their corresponding numerical values.
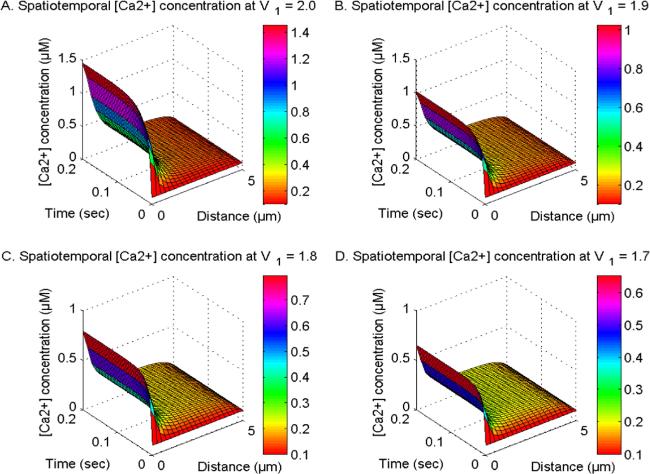
Figure 1 illustrates the spatiotemporal distribution of calcium ion concentration at U1, U2, and U3 = 1.0 and V2 and V3 = 2.0. Additionally, the order of calcium space derivatives (V1) is observed to vary at 2, 1.9, 1.8, and 1.7. Also, this distribution is influenced by the activity of the buffer and several channels, namely the NCX, PMCA, VGCC, Orai channels, and IP3R and ryanodine receptors in neuronal cells. The process of calcium ion diffusion, the expulsion of Ca2+ ions from the cytosol through the SERCA pump, and the fixing of Ca2+ by EGTA buffering mechanisms contribute to a decrease in Ca2+ levels along the spatial dimension. Conversely, the influx of calcium from sources and the increase in Orai channel activity leads to a notable rise in Ca2+ along the temporal dimension over time. The disparity in [Ca2+] diminishes as the fractional space derivatives V1 are reduced from 2 to 1.7 in neurons. The observed range of calcium space derivatives (V1) transitioning from integer-order (2.0) to fractional-order (1.9, 1.8, and 1.7) is linked to the occurrence of superdiffusion events. This phenomenon amplifies the diffusion process and leads to a drop in the spatiotemporal Ca2+ levels. Hence, the phenomenon of superdiffusion exhibited by calcium ions may have a significant impact on mitigating neurotoxicity in disease-related circumstances and maintaining optimal concentration levels.
Figure 1. The spatiotemporal Ca2+ distribution at U1, U2 and U3 = 1.0 and V2V3 = 2.0 with NCX, PMCA, VGCC and Orai channel for V1 = 2, 1.9, 1.8 and 1.7.
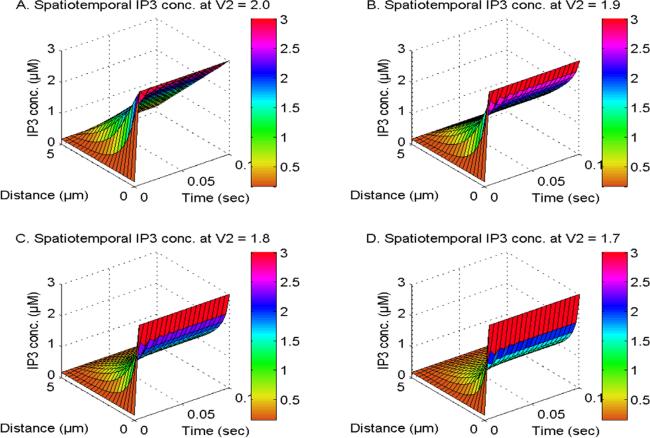
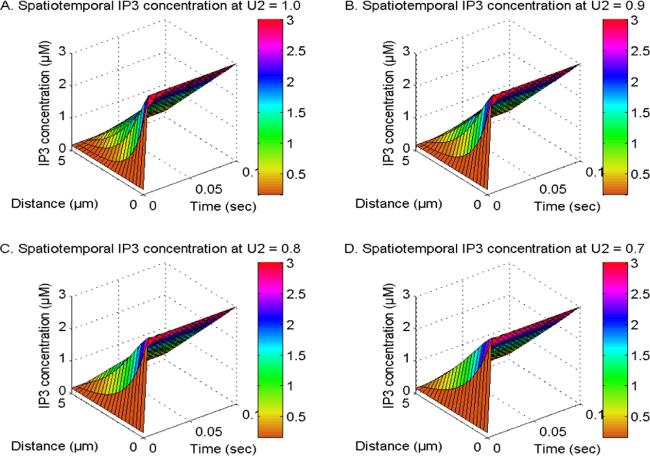
Figure 2 illustrates the spatiotemporal distribution of IP3 at U1, U2 and U3 = 1.0 and V1 and V3 = 2.0. The distribution is observed in the existence of NCX, PMCA, VGCC, and Orai channel, while the order of the IP3 space derivative is considered to be 2, 1.9, 1.8, and 1.7 in neuronal cells. The transportation of inositol trisphosphate molecules and their subsequent breakdown through the actions of kinase and phosphatase enzymes lead to a decrease in the spatial concentration of IP3 in neuronal cells. The proximity of IP3 molecule storage to the IP3-receptor site, which facilitates Ca2+ discharge from the ER to the cytosol, is responsible for the temporal increase in IP3 levels observed in neuronal cells as exhibited in figure 2. The spatiotemporal levels of IP3 molecules exhibit a reduction when the order of space derivative in the IP3 system transitions from an integer-order (V2 = 2.0) to fractional-order (V2 = 1.9, 1.8, and 1.7). Also, the gaps in [IP3] diminish when the order of the spatial derivative decreases from V2 = 2 to 1.7. This reduction can be attributed to the occurrence of superdiffusion events in the IP3 molecules in neuronal cells. The oscillatory behavior of [IP3] is seen due to a discrepancy in the dynamic regulatory mechanisms of IP3 in nerve cells during the early time stages. Hence, the superdiffusion process exhibited by IP3 molecules plays a crucial role in reducing elevated levels of IP3, as well as its related dynamic ions and molecules such as calcium and dopamine in neurons.
Figure 2. The spatiotemporal IP3 distribution at U1, U2 and U3 = 1.0 and V1 and V3 = 2.0 with NCX, PMCA, VGCC and Orai channel for V2 = 2, 1.9, 1.8 and 1.7.
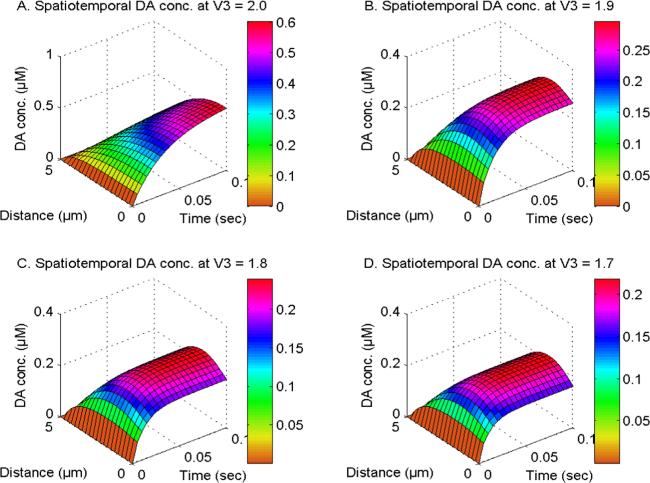
Figure 3 illustrates the spatiotemporal distribution of dopamine at U1, U2 and U3 = 1.0 and V1 and V2 = 2.0. The DA distribution is analyzed in the existence of the NCX, PMCA, VGCC, and Orai channels and the order of the space derivative of the DA system (V3) is varied at 2.0, 1.9, 1.8, and 1.7 in cells. The spatial levels of dopamine are reduced due to the transportation and degradation of DA molecules, whereas the temporal levels of DA in neuronal cells are increased as a result of DA accumulation. It is noted that the levels of dopamine in neurons are regulated by the superdiffusion processes of individual DA molecules. When the order of the DA space derivative lowers from V3 = 2.0 to 1.8, there is a decrease in the concentration of spatiotemporal DA due to the superdiffusion process in neurons. The spatial [DA] exhibits distinct behavior in the integer-order system compared to the fractional-order system. The maximum spatial [DA] is found near the source location for integer-order spatial derivatives. The spatial DA reaches a maximum concentration at the center of neurons for fractional-order space derivatives, attributable to the excessive diffusion of DA closer to the source within neuronal cells. Therefore, the phenomenon of superdiffusion shown by dopamine molecules may play a notable role in reducing the abnormally high levels of DA and DA-dependent ions and molecules in cells under hazardous conditions.
Figure 3. The spatiotemporal DA distribution at U1, U2 and U3 = 1.0 and V1 and V2 = 2.0 with NCX, PMCA, VGCC and Orai channel for V3 = 2, 1.9, 1.8 and 1.7.
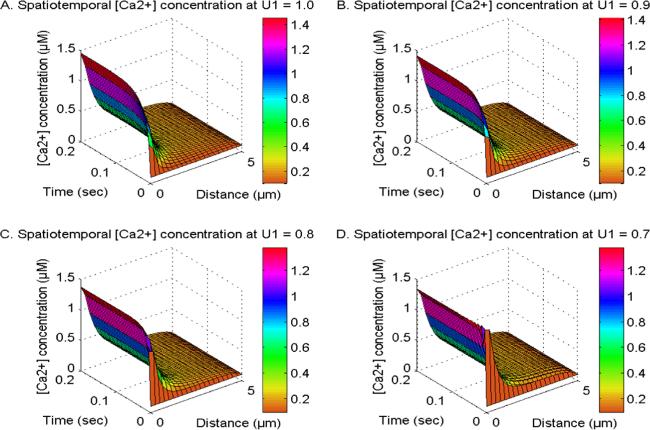
Figure 4 shows the spatiotemporal calcium ion concentration at V1, V2 and V3 = 2.0 and U2 and U3 = 1.0. This Ca2+ distribution is influenced by the active states of NCX, PMCA, VGCC, and Orai channels. The order of time derivative in the Ca2+ system (U1) is evaluated at 1, 0.9, 0.8, and 0.7 in cells. The phenomenon of cell memory exhibiting BM of Ca2+ ions is linked to the temporal order of derivative (0 ≤ U1 ≤ 1) inside cellular systems. When the temporal derivative order transitions from integer (U1 = 1.0) to fractional (0.9, 0.8, and 0.7), the influence of cell memory exhibiting BM of ions on the Ca2+ distribution intensifies in cells. When the value of U1 declines from 1.0 to 0.7, there is a rise in the spatiotemporal Ca2+ levels during the initial period. This elevation is attributed to the cellular memory, which induces Brownian motion effects. Also, the significant consequence of cellular memory exhibiting Brownian motion induces disturbances in calcium signaling, which is characterized by oscillatory patterns in Ca2+ concentration profiles at U1 = 0.7 as illustrated in figure 4(D). As time advances, the temporal calcium levels reach a state of equilibrium concentration for various time derivatives' orders. This occurs because the cellular control mechanism effectively neutralizes the disruptions induced by memory in neuronal cells. Hence, cellular memory may exert adverse impacts on the calcium signaling pathways and their corresponding dynamics such as IP3 and DA within neuronal cells.
Figure 4. The spatiotemporal Ca2+ distribution at V1, V2 and V3 = 2.0 and U2 and U3 = 1.0 with NCX, PMCA, VGCC and Orai channel for U1 = 1, 0.9, 0.8 and 0.7.
Figure 5 illustrates the spatiotemporal distribution of IP3 at V1, V2 and V3 = 2.0 and U1 and U3 = 1.0, considering the existence of PMCA, NCX, VGCC, and Orai channels. The distribution of IP3 is analyzed for varying orders of time derivative in the IP3 system, specifically at U2 values of 1, 0.9, 0.8, and 0.7 in neuronal cells. The fractional order of time derivative in the IP3 system is associated with the memory and BM of IP3 molecules. When the time derivative's order (U2) decreases from an integer value to a fractional value, the elevation in the IP3 concentration is observed during the initial time noted as a result of memory and BM effects, whereas concentration-decreasing processes attempt to lower [IP3] levels in neurons. The discrepancies among these mechanisms induce abnormalities in the distribution of IP3 in the form of fluctuations in neurons. With time, the maintenance of fluctuations in IP3 distribution at appropriate levels in neuronal cells is achieved through an IP3 regulatory mechanism. Thus, the presence of cellular memory of IP3 molecules may contribute to the dysregulation observed in both IP3 and IP3-associated signaling systems like Ca2+ and dopamine.
Figure 5. The spatiotemporal IP3 distribution at V1, V2 and V3 = 2.0 and U1 and U3 = 1.0 with NCX, PMCA, VGCC and Orai channel for U2 = 1, 0.9, 0.8 and 0.7.
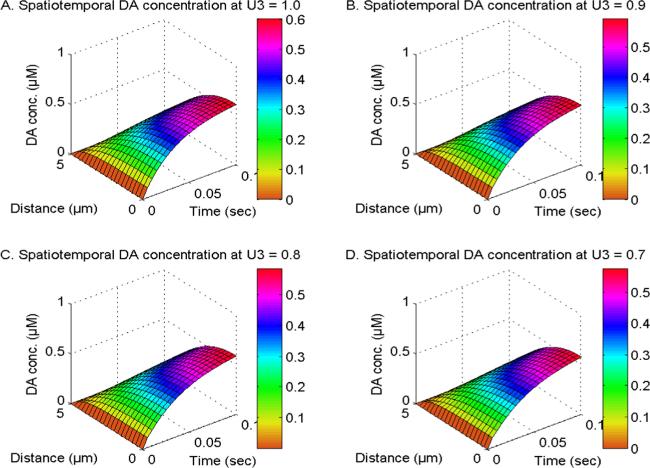
Figure 6 illustrates the spatiotemporal distribution of dopamine (DA) at V1, V2 and V3 = 2.0 and U1 and U3 = 1.0, and the order of DA time derivative (U3) is varied between 1, 0.9, 0.8, and 0.7 in neuronal cells. Also, this distribution is influenced by the presence of NCX, PMCA, VGCC, and Orai channels. When the time derivative of DA reduces from U3 = 1.0 to 0.7, it results in a corresponding decrease in the spatiotemporal DA levels. The influences of the DA cellular memory on the DA distribution differ from the influences of memory of Ca2+ and IP3 on the Ca2+ and IP3 distribution. This is because the DA regulatory mechanism significantly manages the disturbances caused by the cell memory of DA molecules in the initial time. Also, the DA level is dependent on other signaling systems such as calcium and IP3, whose cell memory is not considered. Thus, the decrease in spatiotemporal dopamine (DA) levels resulting from cell memory phenomena has been seen in neuronal cells.
Figure 6. The spatiotemporal DA distribution at V1, V2 and V3 = 2.0 and U1 and U2 = 1.0 with NCX, PMCA, VGCC and Orai channel for U3 = 1, 0.9, 0.8 and 0.7.
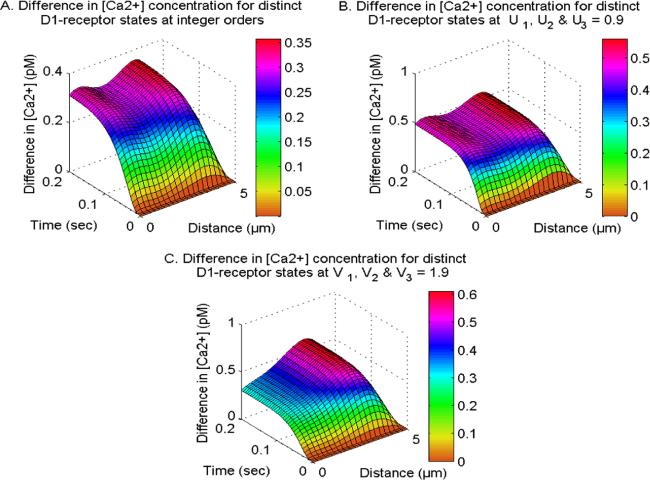
Figure 7 depicts the effects of the D1-receptor on the calcium concentration profiles at different integer-order and fractional-order in neuron cells. The activation of the D1-receptor stimulates the synthesis of IP3 molecules, which in turn amplifies the calcium concentration by activating IP3R in neurons. Deactivating the D1-receptor decreases the production of IP3, causing a subsequent decrease in Ca2+ levels in cells. The difference in the Ca2+ levels for distinct D1-receptor states exhibits different behavior at integer and fractional order systems within neurons. The functioning of the D1-receptor is contingent upon the calcium and DA levels within neurons. The modulation of D1-receptor is directly correlated with dopamine levels and inversely correlated with calcium levels in neurons, which further causes uneven distribution of calcium concerning distinct D1-receptor states within neurons. The functioning of the D1-receptor enhances at the fractional-order temporal and spatial derivatives due to memory and superdiffusion processes. Hence, the D1-receptor significantly influences the allocation of Ca2+ across various integer and fractional-order derivatives. Additionally, it exhibits a feedback mechanism that involves Ca2+, IP3, and DA in neurons.
Figure 7. The difference in spatiotemporal Ca2+ distribution at integer and fractional orders with NCX, PMCA, VGCC and Orai channel for distinct D1-receptor states.
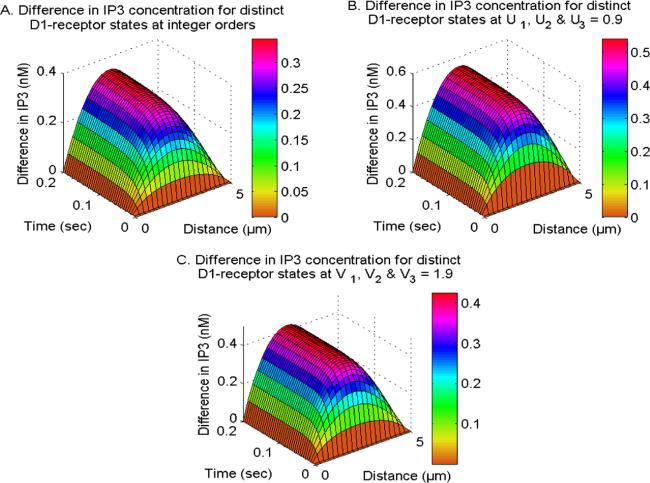
Figure 8 illustrates the impact of the active and inactive D1-receptors phases on the spatiotemporal neuronal IP3 levels. The presence of the D1-receptor increases the creation of IP3 molecules, while the absence of the D1-receptor decreases the generation of IP3 in neurons. The spatial gradient in IP3 levels between the active and inactive phases of the D1-receptor rises from x = 0 μm towards the center of cells and subsequently decreases towards the other end (x = 5 μm) since the appropriate IP3 concentration levels are maintained at both the boundaries within neurons. The temporal disparities in IP3 concentrations between active and inactive D1-receptor phases increase throughout time and reach a state of balance at various locations within neurons. The difference in IP3 concentration levels increases when the fractional time derivatives (U1, U2 and U3 = 0.9) are considered due to the memory effects of ions and molecules within neurons. Furthermore, the molecules exhibit superdiffusion, which leads to a reduction in the differences in IP3 concentration levels. This effect is observed when the fraction space derivatives (V1, V2 and V3 = 1.9) are taken into account. Therefore, the D1-receptor has a substantial impact on the distribution of IP3 at different integer as well as fractional-order derivatives, and it also demonstrates a feedback loop involving Ca2+, IP3, and DA in neurons.
Figure 8. The difference in spatiotemporal IP3 distribution at integer and fractional orders with NCX, PMCA, VGCC and Orai channel for distinct D1-receptor states.
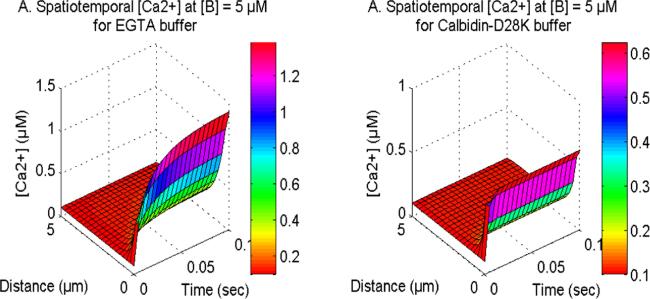
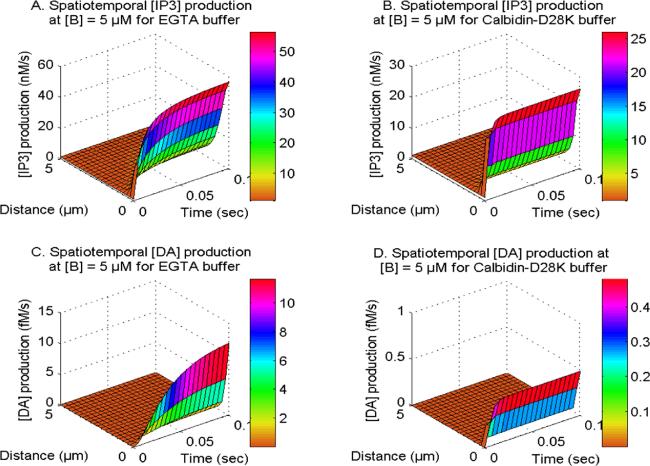
Figure 9 displays the spatiotemporal Ca2+ concentration for distinct buffers namely EGTA and Calbindin-D28K in neurons. The buffer molecule exhibits an affinity to the excess cytosolic Ca2+ ions to bind them, hence reducing the cytosolic Ca2+ within cells. Upon analysis of the distinct buffers, it is evident that neuronal cells have a higher reduction in calcium levels for the Calbindin-D28K buffer as compared to the EGTA buffer. The Calbindin-D28K buffer has a higher affinity for binding calcium ions in contrast to the EGTA buffer, which results in the temporal [Ca2+] profile reaching steady-state more rapidly than the EGTA buffer within the cell. Similarly in figure 10, the spatiotemporal IP3 and DA production fluxes, which are reliant on Ca2+ levels, also demonstrate a comparable pattern for EGTA and Calbindin-D28K buffers in neurons. In the case of Calbindin-D28K buffer, there is a decreased amount of IP3 and DA production in cells. Hence, the Calbindin-D28K buffer plays a significant role in maintaining the appropriate levels of calcium, IP3, and DA in situations involving neurotoxicity and illness.
Figure 10. The spatiotemporal IP3 and DA production with NCX, PMCA, VGCC and Orai channel for EGTA and Calbidin-D28K buffers.
Table 2 displays the absolute relative approximate errors for Ca2+, IP3, and DA concerning GSI. At GSI = 100, the lowest accuracy and maximum error of the model for Ca2+, IP3, and DA are 99.9920567% and 0.0079433%, respectively.
Table 2. Absolute relative error approximations for [Ca2+], [IP3] and [DA] distribution.
GSI
Errors for [Ca2+]
Errors for [IP3]
Errors for [DA]
0 μm
0.25 μm
0 μm
10 and 11
0.053521%
0.88%
12.20%
20 and 21
0.0055890%
0.29%
6.67%
50 and 51
1.9631 × 10−4%
0.011438%
0.72%
100 and 101
7.5399 × 10−7%
4.4128 × 10−5%
0.0079433%
The numerical results of the integer-order Ca2+ and IP3 systems are displayed in table 3. These findings are consistent with the documented results [42] at a time of 50 s, indicating a good agreement between the current findings and existing research. The root mean square errors for Ca2+ and IP3 were determined to be 0.0035 and 0.000 119 95, sequentially. Nevertheless, the lack of experimental documentation on the kinetics of fractional-order calcium and IP3 is a significant challenge in terms of attaining thorough validation. Also, the numerical outcomes related to the fractional concentration of calcium ions, IP3, and dopamine signaling mechanisms within neuronal cells exhibit conformity with biophysical realities.
Table 3. [Ca2+] and [IP3] at t = 50 s compared to Wagner et al [42].
Tables 4 and 5 depict the comparison in the Ca2+, IP3 and DA levels (in μM) among three interacting systems: Ca2+, IP3 and DA, with two interacting systems including Ca2+ and IP3 and Ca2+ and DA at distinct neuronal sites. The previous fractional-order model of the two signaling systems of Ca2+ and IP3 included the bi-directional feedback between calcium and IP3 in neurons. However, the influence of the ryanodine receptor on the interacting calcium and IP3 was not addressed in the prior fractional-order model. The dynamics of fractional-order calcium and dopamine, together with the effects of the ryanodine receptor, were included; however, there is solely unidirectional feedback between calcium and dopamine in neurons. The present study establishes the feedback loop mechanism among fractional-order Ca2+, IP3, and DA signaling systems by integrating the D1-receptor in the regulation of IP3 in neurons. The present work incorporates various essential cellular processes namely the PMCA channel, Orai channel, VGCC, and NCX, which control Ca2+, IP3, and DA at optimal levels in neurons. The impacts of these pathways were not present in the previously reported research. Consequently, the interplay of Ca2+, IP3, and DA signaling pathways, in conjunction with biological mechanisms such as the D1-receptor, PMCA channel, Orai channel, RyR, VGCC, and NCX, renders the current study more advanced than previously published research.
Table 4. Comparison of [Ca2+] in three signaling systems of Ca2+, IP3, and DA with the [Ca2+] in two interacting systems of Ca2+ and IP3 and Ca2+ and DA.
Locations (in μm)
[Ca2+] for two signaling systems; Ca2+ and IP3
[Ca2+] for two signaling systems; Ca2+ and DA
[Ca2+] for three signaling systems; Ca2+, IP3, and DA
0
1.59352626311615
1.67374166702743
1.39639145777306
1
0.959594060751098
1.10734892213237
0.444646823496717
2
0.583964456594164
0.775884667935086
0.199507243264428
3
0.360415236709176
0.573624721540783
0.152937005272446
4
0.212589240812298
0.377880288715064
0.136796466146167
5
0.1
0.1
0.1
Table 5. Comparison of [IP3] and [DA] in three signaling systems of Ca2+, IP3, and DA with the [IP3] and [DA] in two interacting systems of Ca2+ and IP3 and Ca2+ and DA.
Locations (in μm)
IP3 for two signaling systems; Ca2+ and IP3
IP3 for three signaling systems; Ca2+, IP3, and DA
DA for two signaling systems; Ca2+ and DA
DA for three signaling systems; Ca2+, IP3, and DA
0
3
3
0.633949375794030
0.634047185204713
1
2.43213661177129
2.43164738874235
0.611019641480479
0.611104126486061
2
1.86430790558617
1.86361274208202
0.538408754743416
0.538474355261389
3
1.29639654811965
1.29574921058797
0.412791833315504
0.412836720211876
4
0.728322817076785
0.727927224597782
0.233790129029834
0.233813079757628
5
0.16
0.16
0
0
Conclusion
A mathematical model has been framed for interactive systems of nonlinear calcium, IP3, and dopamine, utilizing a fractional-order approach along spatial and temporal dimensions. The CN method with the Grunwald technique for fractional spatial derivatives and the L1 scheme for fractional temporal derivatives in combination with GSI were employed to get numerical findings. The L1 scheme is exceptionally resilient and effective for the analysis of fractional-order differential equations. The fundamental characteristic of the L1 scheme is its capacity to integrate a memory trail that records and assimilates all previous occurrences, essential for simulating the dynamics. Earlier researchers examined the fractional-order reaction-diffusion models of calcium and DA incorporating unidirectional feedback in neurons. However, no fractional-order model has been documented about the bi-directional feedback process between calcium and dopamine in neuronal cells. The influence of DA on calcium signaling is facilitated by the integration of IP3 signaling molecules, as DA signaling plays a crucial role in the synthesis of IP3 molecules. The interplay of IP3 signaling with calcium and the dopamine system generates a feedback loop among these three dynamic systems. No study is presently documented on the interacting fractional-order systems of Ca2+, IP3 and DA in neuronal cells. Also, the current model offers novel viewpoints on the implications of superdiffusion processes and cellular memory, which exhibit BM on the cooperation of fractional-order nonlinear dynamics of Ca2+, IP3, and DA in neurons. To assess the model's performance, the absolute relative approximation errors for Ca2+, IP3, and DA concentrations in neurons at various sites have been computed across multiple GSI. At 100 iterations, the model's minimum accuracy and maximum error for Ca2+, IP3, and DA are recorded at 99.9920567% and 0.0079433%, respectively. The spectral radius has been calculated to assess stability and is found to be less than 1, indicating the unconditional stability of the numerical techniques applied.
From the aforementioned results, the following basic conclusions can be drawn:
1. The superdiffusion phenomenon is a concentration-reducing mechanism. Enhanced diffusion promotes a more comprehensive and efficient reduction of intracellular Ca2+ levels through improved accessibility to binding sites and buffers, effective removal by intracellular organelles, membrane extrusion mechanisms, and feedback regulation.
2. Cellular memory induces an increase in the concentration of ions and molecules during the early period. Over time, it begins to diminish the concentration levels of ions and molecules within neurons.
3. Various buffer types, specifically EGTA and Calbindin-D28K, diminish cytosolic calcium ion concentrations in differing amounts due to their distinct interaction rates with calcium in neurons.
The obtained numerical results have led to the following novel conclusions:
1. A significant decrease in the spatiotemporal concentration levels of Ca2+, IP3, and dopamine has been observed due to the superdiffusion mechanism in neuronal cells. In conditions associated with disease and characterized by high concentrations, the cell may use superdiffusion phenomena as an approach for reducing neurotoxicity. Any disturbances in the superdiffusion process can result in increased levels of Ca2+, IP3, and dopamine, leading to the disruption of the interactive spatiotemporal dynamical system involving these ions and molecules.
2. The impact of DA cell memory on DA concentration levels varies from the impact of calcium and IP3 memory on calcium and IP3 concentration in neurons. The reason for this behavior can be attributed to the regulatory mechanism of dopamine, which effectively controls the disruptions induced by cell memory and Brownian motion of DA molecules within neuronal cells during initial time.
3. The D1-receptor influences the concentration of calcium, IP3, and DA in neurons. The functioning of D1-receptor at fractional-order space and time derivatives significantly differs from the integer-order space and time derivatives within neurons due to the memory and accelerated diffusion phenomena and the same is visible in our findings.
4. The neuronal cells experience a greater decrease in calcium levels concerning Calbindin-D28K buffer compared to the EGTA buffer, which further leads to reduced IP3 and DA synthesis in neuronal cells.
Therefore, it may be inferred that the phenomenon of super-diffusion and memory which involves the BM of calcium, IP3, and DA have crucial functions in governing the dynamical signaling systems of neuronal Ca2+, IP3 and dopamine. The deregulation of any or all of these pathways can result in neurotoxic circumstances and contribute to the progression of disease-associated conditions such as AD.
The interaction of three signaling ions and molecules namely calcium, IP3, and DA has been considered in the proposed model. However, the current study establishes a foundation for future investigations into the dynamic interactions of critical neuronal signaling molecules, including Aβ, PLC, and ATP, with nonlinear calcium, IP3, and DA dynamics in both healthy and pathological neuronal states. Moreover, the intricate connections of additional essential signaling molecules with the calcium, IP3, and DA systems will provide a more precise comprehension of several illnesses, including Alzheimer's disease and Parkinson's disease.
The CN approach, Grunwald and L1 schemes, and GS iterations have demonstrated significant efficacy in obtaining essential insights into the interactive Ca2+, IP3, and DA signaling systems within neuronal cells. This study provides insights into the roles of superdiffusion and cellular memory signaling molecules, as well as the essential signaling mechanisms regulating the concentration levels of calcium, IP3, and DA in neurons. Also, the information acquired through these models on the basic mechanisms and events linked with disease-related states can be valuable to biomedical researchers in the advancement of diagnostic and therapeutic interventions.
Appendix. Model equations summary
The fractional derivatives along space in equations (1), (14) and (20) exhibit the utilization of the CN method in conjunction with the Grunwald formula as follows [78],
Where, the h denotes the spatial increment for i = 1, 2, 3,…, K − 1. The normalized Grunwald weights referred to as g1k, g2k, and g3k, are contingent upon the index k and the orders V1, V2, and V3, in a sequential manner. The representation of these weights is as follows:
Defining, $B1={D}_{{\rm{Ca}}}\tfrac{{\rm{\Gamma }}\left(2-{U}_{1}\right){\left({\rm{\Delta }}t\right)}^{{U}_{1}}}{2{h}^{{V}_{1}}}$, then equation (47) can be expressed in the following manner
where $B3={D}_{\mathrm{DA}}\tfrac{{\rm{\Gamma }}(2-{U}_{3}){({\rm{\Delta }}t)}^{{U}_{3}}}{2{h}^{{V}_{3}}}.$
Consider λ1 is an eigenvalue of A1, such that A1X = λ1X for some non-zero vector X. Let $\left|{x}_{i}\right|\,=\max \left\{\left|{x}_{j}\right|:j={\rm{0}},{\rm{1}},{\rm{2}},\mathrm{..}.,K\right\}$, then $\displaystyle {\sum }_{j=0}^{K}A{1}_{i,j}{x}_{j}={{\rm{\lambda }}}_{1}{x}_{i}$, this implies
Since, $\left(\displaystyle {\sum }_{K=0}^{\infty }g{1}_{K}=0\right)$ and g11 is the sole element in the Grunwald weights sequence that possesses a negative value, and $g{1}_{1}=-{V}_{1}$ and thus for ${1\lt V}_{1}\leqslant {\rm{2}}$
Since $\left|\tfrac{{x}_{j}}{{x}_{i}}\right|\leqslant 1\,\mathrm{and}\,g{1}_{{\rm{j}}}\geqslant 0,\mathrm{for}\,j={\rm{0}},{\rm{2}},{\rm{3}},\mathrm{..}.,$
Given that B1, B2, and B3 represent non-negative real numbers, all eigenvalues of A1, A2, A3, and A satisfy the criterion $\left|{\rm{\lambda }}\right|\geqslant 1$. Matrix A is considered to be invertible, and it is observed that every eigenvalue of the inverse matrix A−1 meets a condition $\left|\eta \right|\leqslant 1$. Hence, it can be concluded that the spectral radius of the matrix $\rho \left({A}^{\,-1}\right)\leqslant 1$, which implies that the used scheme exhibits unconditional stability.
PalopJ J, JonesB, KekoniusL, ChinJ, YuG Q, RaberJ, MasliahE, MuckeL2003 Neuronal depletion of calcium-dependent proteins in the dentate gyrus is tightly linked to Alzheimer's disease-related cognitive deficits Proc. Natl. Acad. Sci. USA100 9572 9577
TuH, NelsonO, BezprozvannyA, WangZ, LeeS F, HaoY H, SerneelsL, De StrooperB, YuG, BezprozvannyI2006 Presenilins form ER Ca2+ leak channels, a function disrupted by familial Alzheimer's disease-linked mutations Cell126 981 993
TewariS G, PardasaniK R2010 Finite element model to study two dimensional unsteady state cytosolic calcium diffusion in presence of excess buffers IAENG Int. J. Appl. Math.40 1 5
12
TewariS G, PardasaniK R2012 Modeling effect of sodium pump on calcium oscillations in neuron cells J. Multiscale Model.04 1250010
TripathiA, AdlakhaN2011 Closed form solution to problem of calcium diffusion in cylindrical shaped neuron cell World Acad. Sci. Eng. Technol.80 739 742
14
JhaA, AdlakhaN2014 Analytical solution of two dimensional unsteady state problem of calcium diffusion in a neuron cell J. Med. Imaging Heal. Inform.4 547 553
PawarA, PardasaniK R2022 Simulation of disturbances in interdependent calcium and β-amyloid dynamics in the nerve cell Eur. Phys. J. Plus 2022 1378 137 1 23
JhaB K, AdlakhaN, MehtaM N2013 Two-dimensional finite element model to study calcium distribution in astrocytes in presence of VGCC and excess buffer Int. J. Model. Simulation, Sci. Comput.4 1250030
JhaB K, AdlakhaN, MehtaM N2014 Two-dimensional finite element model to study calcium distribution in astrocytes in presence of excess buffer Int. J. Biomath.7 1 11
PathakK, AdlakhaN2016 Finite element model to study two dimensional unsteady state calcium distribution in cardiac myocytes Alexandria J. Med.52 261 268
PathakK B, AdlakhaN2015 Finite element model to study calcium signalling in cardiac myocytes involving pump, leak and excess buffer J. Med. Imaging Heal. Inform.5 683 688
PandayS, PardasaniK R2013 Finite element model to study effect of advection diffusion and Na +/Ca2+ exchanger on Ca2+ distribution in oocytes J. Med. Imaging Heal. Inform.3 374 379
NaikP A, PardasaniK R2019 Three-dimensional finite element model to study effect of RyR calcium channel, ER leak and SERCA pump on calcium distribution in oocyte cell Int. J. Comput. Methods16 1 19
NaikP A, PardasaniK R2016 Finite element model to study calcium distribution in oocytes involving voltage gated Ca2+ channel, ryanodine receptor and buffers Alexandria J. Med.52 43 49
NaikP A, PardasaniK R2015 One dimensional finite element model to study calcium distribution in oocytes in presence of VGCC, RyR and buffers J. Med. Imaging Heal. Inform.5 471 476
NaikP A, PardasaniK R2017 Three-dimensional finite element model to study calcium distribution in oocytes Netw. Model. Anal. Heal. Inform. Bioinforma.6 1 11
KotwaniM, AdlakhaN, MehtaM N2014 Finite element model to study the effect of buffers, source amplitude and source geometry on spatiooral calcium distribution in fibroblast cell J. Med. Imaging Heal. Inform.4 840 847
JagtapY, AdlakhaN2018 Finite volume simulation of two dimensional calcium dynamics in a hepatocyte cell involving buffers and fluxes Commun. Math. Biol. Neurosci.2018 1 16
31
MishraV, AdlakhaN2023 Numerical simulation of calcium dynamics dependent ATP degradation, IP3 and NADH production due to obesity in a hepatocyte cell J. Biol. Phys.49 415 442
MishraV, AdlakhaN2023 Spatio temporal interdependent calcium and buffer dynamics regulating DAG in a hepatocyte cell due to obesity J. Bioenerg. Biomembr.55 249 266
BhardwajH, AdlakhaN2022 Radial basis function based differential quadrature approach to study reaction diffusion of Ca2+ in T Lymphocyte Int. J. Comput. Methods20 2250059
TewariV, TewariS, PardasaniK R2011 A model to study the effect of excess buffers and Na+ ions on Ca2+ diffusion in neuron cell World Acad. Sci. Eng. Technol.76 41 46
JhaA, AdlakhaN2015 Two-dimensional finite element model to study unsteady state Ca2+ diffusion in neuron involving ER LEAK and SERCA Int. J. Biomath.8 1550002
FalckeM, HuertaR, RabinovichM I, AbarbanelH D I, ElsonR C, SelverstonA I2000 Modeling observed chaotic oscillations in bursting neurons: the role of calcium dynamics and IP3Biol. Cybern.82 517 527
YoungG W D E, KeizerJ1992 A single-pool inositol 1,4,5-trisphosphate-receptor-based model for agonist-stimulated oscillations in Ca2+ concentration Biophysics89 9895 9899
LiY X, RinzelJ1994 Equations for InsP3 receptor-mediated [Ca2+]i oscillations derived from a detailed kinetic model: a Hodgkin–Huxley like formalism J. Theor. Biol.166 461 473
WagnerJ, FallC F, HongF, SimsC E, AllbrittonN L, FontanillaR A, MoraruI I, LoewL M, NuccitelliR2004 A wave of IP3 production accompanies the fertilization Ca2+ wave in the egg of the frog, Xenopus laevis: theoretical and experimental support Cell Calcium35 433 447
SinghN, AdlakhaN2019 A mathematical model for interdependent calcium and inositol 1,4,5-trisphosphate in cardiac myocyte Netw. Model. Anal. Heal. Inform. Bioinforma.8 18
JagtapY, AdlakhaN2023 Numerical model of hepatic glycogen phosphorylase regulation by nonlinear interdependent dynamics of calcium and IP3Eur. Phys. J. Plus138 399
Vaishali, AdlakhaN2023 Disturbances in system dynamics of Ca2+ and IP3 perturbing insulin secretion in a pancreatic β-cell due to type-2 diabetes J. Bioenerg. Biomembr.55 151 167
KothiyaA B, AdlakhaN2023 Cellular nitric oxide synthesis is affected by disorders in the interdependent Ca2+ and IP3 dynamics during cystic fibrosis disease J. Biol. Phys.49 133 158
BhardwajH, AdlakhaN2024 Model to study interdependent calcium and IP3 distribution regulating NFAT production in T Lymphocyte J. Mech. Med. Biol.24 2350055
PawarA, PardasaniK R2022 Effects of disorders in interdependent calcium and IP3 dynamics on nitric oxide production in a neuron cell Eur. Phys. J. Plus137 543
PawarA, PardasaniK R2023 Mechanistic insights of neuronal calcium and IP3 signaling system regulating ATP release during ischemia in progression of Alzheimer's disease Eur. Biophys. J.
SodenM E, JonesG L, SanfordC A, ChungA S, GülerA D, ChavkinC, LujánR, ZweifelL S2013 Disruption of dopamine neuron activity pattern regulation through selective expression of a human KCNN3 mutation Neuron80 997 1009
MosharovE V, LarsenK E, KanterE, PhillipsK A, WilsonK, SchmitzY, KrantzD E, KobayashiK, EdwardsR H, SulzerD2009 Article interplay between cytosolic dopamine, calcium, and α-synuclein causes selective death of substantia nigra neurons Neuron62 218 229
PawarA, PardasaniK R2023 Computational model of calcium dynamics-dependent dopamine regulation and dysregulation in a dopaminergic neuron cell Eur. Phys. J. Plus123 1 19
NaikP A, YeolekarB M, QureshiS, YeolekarM, MadzvamuseA2024 Modeling and analysis of the fractional-order epidemic model to investigate mutual influence in HIV/HCV co-infection Nonlinear Dyn.112 11679 11710
JoshiH, JhaB K2022 2D dynamic analysis of the disturbances in the calcium neuronal model and its implications in neurodegenerative disease Cogn. Neurodyn.17 1637 1648
EzzatM A, KaramanyA S E2011 Fractional order heat conduction law in magneto-thermoelasticity involving two temperatures Z. Angew. Math. Phys.62 937 952
PawarA, PardasaniK R2023 Fractional-order reaction—diffusion model to study the dysregulatory impacts of superdiffusion and memory on neuronal calcium and IP3 dynamics Eur. Phys. J. Plus138 780
PawarA, PardasaniK R2024 Nonlinear system dynamics of calcium and nitric oxide due to cell memory and superdiffusion in neurons Commun. Theor. Phys.76 055002
SimsC E, AllbrittontN L1998 Metabolism of inositol 1,4,5-trisphosphate and inositol 1,3,4,5- tetrakisphosphate by the oocytes of Xenopus laevis J. Biol. Chem.273 4052 4058
BugrimA, FontanillaR, EutenierB B, KeizerJ, NuccitelliR2003 Sperm initiate a Ca2+ wave in frog eggs that is more similar to Ca2+ waves initiated by IP3 than by Ca2+Biophys. J.84 1580 1590
TadjeranC, MeerschaertM M, SchefflerH P2006 A second-order accurate numerical approximation for the fractional diffusion equation J. Comput. Phys.213 205 213




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}