



1. Introduction
The mechanical properties of membranes play a crucial role in their function, structure, and dynamics in biological systems [1]. These properties are primarily governed by the composition and structure of the lipid bilayer, and its interactions with proteins. In 1973, by making analogies between the lipid bilayer and the liquid crystal in their orientational order, Helfrich proposed an elastic theory for the cell membrane [2]. As the thickness of the membrane is much smaller than its lateral dimension, the theory assumes that the membrane is a two-dimensional surface embedded in three-dimensional space, and deforming under forces while maintaining its in-plane fluidity [2,3]. By writing the bending energy of a membrane in terms of the mean curvature and the Gaussian curvature of the membrane surface, the Helfrich theory has been successfully applied to describing and predicting various deformation phenomena of cell membranes, including the biconcave shape of the red blood cells [4,5], vesicle shape transformation [6-11], membrane fusion and fission [12-15], as well as membrane-protein interaction [16-22].
In Helfrich theory, the mechanical properties of the membrane are characterized by three phenomenological parameters: the bending modulus κ, the Gaussian curvature modulus $\bar{\kappa }$, and the spontaneous curvature c0. The equilibrium membrane shape is obtained by minimizing the elastic energy subject to geometric constraints. The bending modulus κ quantifies the energetic cost of deforming the membrane's mean curvature from its spontaneous curvature c0. Any deformation that induces membrane curvature could be related to κ, therefore, experimental determination of the bending modulus κ can be achieved by probing the mechanical response of the membrane to forces [23-27] or thermal fluctuations [28-33]. For instance, measuring the force required to pull a membrane tether from a giant vesicle [25-27] or the fluctuation spectrum of a freely standing membrane [31-33] is widely used to obtain the value of the bending modulus.
Spontaneous curvature c0 reflects the membrane's tendency to adopt a preferred curvature even in the absence of external forces. Any types of asymmetry between the two leaflets, such as the lipid composition, lipid number density, and protein occupation, could induce spontaneous curvature [34-37]. Directly measuring the spontaneous curvature of a membrane in experiments is challenging, as its effect is always intertwined with the bending modulus κ and the membrane tension. However, a comparison between the experimentally observed membrane shape and the theory-predicted membrane shape in response to an asymmetric factor provides an indirect approach to extract the spontaneous curvature. For instance, in simulations, the spontaneous curvature has been obtained by the axial force of the tubular film and the average curvature of the curved film strip [38].
Unlike the bending modulus κ governing smooth deformations, the Gaussian bending modulus $\bar{\kappa }$ contributes to the total energy only in processes involving changes in topology (e.g., transitions from a sphere to a torus, formation of pores [39], vesicle fusion and fission [40-44], endocytosis and exocytosis [45]) due to the famous Gauss-Bonnet theorem [46,47]. This makes direct measurement of $\bar{\kappa }$ less straightforward. As the formation and closure of membrane pores involve Gaussian curvature contributions [39], measuring the transition probability from a curved strip to a closed vesicle [48,49] or reading the fluctuations at the edge of an open membrane [50] has been used to determine the Gaussian curvature modulus $\bar{\kappa }$ in molecular dynamics simulations. As the fusion pore structure formed in the fusion process is sensitive to the Gaussian curvature modulus [42], experiments on the bicontinuous inverted cubic phases of amphiphiles can accurately determine the value of the Gaussian curvature modulus [44,51,52].
Meanwhile, in molecular dynamics simulations, the spontaneous curvature can be determined from the first moment of the lateral stress profile [53], while the Gaussian curvature modulus can be obtained from the second moment [54], even though discrepancies have been reported between this stress-profile-based method and the shape-based method [55]. These differences can be attributed to the omission of the variance term in the derivation of the stress-profile method [56].
The buckling phenomenon is a mechanical instability that occurs when a structure subject to compressive stress undergoes a sudden deformation or collapse [57]. In various biological processes, such as the spiky shape of red blood cells induced by osmotic stress [58,59], or the formation of filopodia during cell migration [60], such membrane buckling is associated with a creation of surface folds or wrinkles. Here it should be noted that buckling of a fluid membrane exhibits different behaviors compared with that of a solid sheet, such as anisotropic stress and negative compressibilities [61,62]. This has been used as a robust method to extract the bending modulus κ of a rectangular membrane when subject to compressive stresses [62].
In our previous work, we proposed a method to measure the Gaussian curvature modulus $\bar{\kappa }$ of a circular membrane via a buckling protocol [63]. The membrane is subject to radially oriented compressive stresses and the critical stress to buckled the membrane is found to be dependent on $\bar{\kappa }$ if no external torque is applied at the edge of the membrane. However, the effect of the spontaneous curvature is not considered in this study. In this work, we extend our previous work to conditions where the buckled membrane possesses a nonzero spontaneous curvature. It is shown that the stress-strain relationship of a buckled membrane depends on both the Gaussian curvature modulus $\bar{\kappa }$ and the spontaneous curvature c0. Based on this observation, we propose a method capable of measuring $\bar{\kappa }$ and c0 simultaneously.
2. Theoretical model
The membrane in this paper is modelled as a two-dimensional axisymmetric surface parameterized as
$\begin{eqnarray}{\boldsymbol{X}}(u,\phi )=[r(u)\cos \phi ,r(u)\sin \phi ,z(u)],\end{eqnarray}$
where the parameter Φ ∈ [0, 2π] and u ∈ [0, 1], with u=0 and u=1 corresponding to the membrane tip and the membrane base, respectively (figure 1). The shape functions r(u) and z(u) satisfy $\begin{eqnarray}{r}^{{\prime} }=h\cos \psi ,\quad \quad \,{z}^{{\prime} }=-h\sin \psi ,\end{eqnarray}$
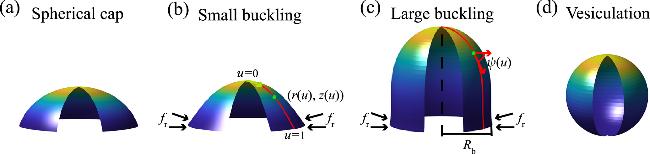
Figure 1. Illustration of deformed circular membranes with (a) a spherical cap, (b) a small buckling, and (c) a large buckling or (d) a vesiculation under free-hinge BC, considering spontaneous curvature. |
where ψ(u) represents the angle spanned between the tangential direction of the meridian curve eu=∂X/∂u and the radial direction ${{\boldsymbol{e}}}_{r}=[\cos \phi ,\sin \phi ,0]$, h(u) is a scaling factor that fulfills the equation $h=\sqrt{{r{}^{{\prime} }}^{2}+{z{}^{{\prime} }}^{2}}$. Hereafter, we use the prime to denote the derivative with respect to u. Note that when the parameter u varies from 0 to 1, the two coordinate expressions [r(u), z(u)] and [r(u2), z(u2)] describe the same membrane shape but differ in the expression of h. Due to this redundancy, we impose ${h}^{{\prime} }=0$, indicating that h is a constant, which is equal to the total arclength of the membrane profile.
For a membrane with a spontaneous curvature c0, its bending energy can be written as
$\begin{eqnarray}{F}_{\,\rm{b}\,}={\int }_{0}^{1}\left[\frac{\kappa }{2}{({c}_{1}+{c}_{2}-{c}_{0})}^{2}+\bar{\kappa }{c}_{1}{c}_{2}\right]2\pi rh\,\rm{d}\,u,\end{eqnarray}$
where κ and $\bar{\kappa }$ denote the bending rigidity of the membrane associated with the mean curvature and the Gaussian curvature, respectively. the The two principal curvatures of the membrane surface are given by ${c}_{1}={\psi }^{{\prime} }/h$ and ${c}_{2}=(\sin \psi )/r$. We assume that the membrane is in a homogeneous state such that the three parameters κ, $\bar{\kappa }$ and c0 are constant.To account for the compressive stress exerted at the membrane base, we introduce a boundary energy term C . The total free energy F of the membrane includes the bending energy and the boundary energy, 2 ). Variations of the functional $\bar{F}$ with respect to the shape variables lead to shape equations (A3 ), (A4 ) and (A5 ), as well as boundary conditions (BCs) (B1 )-(B6 ) and (B9 ), together constituting a boundary value problem (see appendix A and appendix B for the mathematical details). In general, there are two types of BCs, depending on whether the angle at the membrane base is free to rotate (free-hinge BC) or fixed to a constant value (fixed-hinge BC). We have shown previously that only with the free-hinge BC will the buckling process depend on the Gaussian bending rigidity $\bar{\kappa }$ [63], therefore, in this study we restrain our investigation to the free-hinge BC, which is given by
$\begin{eqnarray}{F}_{\rm{l}\,}={f}_{\rm{r}}\pi {R}_{\,\rm{b}}^{2},\end{eqnarray}$
where fr denotes the magnitude of the compressive stress pointing inwards along the radial direction, and Rb denotes the radius of the membrane base. Here we neglect the energy contribution made by the edge tension γ which is proportional to the perimeter of the boundary circle, 2πRbγ [39,64,68]. The possible effect caused by γ is discussed in appendix $\begin{eqnarray}F={F}_{\rm{b}\,}+{F}_{\,\rm{l}}.\end{eqnarray}$
We assume that the membrane is incompressible, which means that the total surface area A0 of the membrane is conserved when deformed. Such a constraint is imposed by introducing a Lagrangian multiplier σ conjugated to the surface area A, $\begin{eqnarray}{F}_{\sigma }=\sigma A.\end{eqnarray}$
The effective energy of the membrane $\bar{F}=F+{F}_{\sigma }$ can be expressed as a functional of the shape variables for the meridian profile: $\begin{eqnarray}\bar{F}=2\pi \kappa {\int }_{0}^{1}{ \mathcal L }\left[\psi ,{\psi }^{{\prime} },r,{r}^{{\prime} },z,{z}^{{\prime} },h,\gamma ,\eta \right]\,\rm{d}\,u,\end{eqnarray}$
where γ and η are Lagrangian multipliers that enforce the geometric constraints in equation ( $\begin{eqnarray}\left[\kappa ({c}_{1}+{c}_{2}-{c}_{0})+\bar{\kappa }{c}_{2}\right]{| }_{u=1}=0.\end{eqnarray}$
Physically, it implies torque-free at the edge of the membrane and has been widely used for studying membranes with an open edge [39,64], and the problem of pulling a membrane tube from a vesicle [65].In the absence of boundary stresses, we find a spherical cap solution that satisfies all the shape equations (A3 )-(A5 ) with the associated BCs (B1 )-(B6 ), and (B9 ) (see appendix E ). The radius of curvature of the spherical cap reads 8 ), given that a spherical cap has c1=c2=1/Rcap (figure 1(a)). The spanning angle θ of the spherical cap is determined by its surface area A0 via the equation
$\begin{eqnarray}{R}_{{\rm{cap}}}=\frac{2+\bar{\kappa }/\kappa }{{c}_{0}},\end{eqnarray}$
which is obtained from equation ( $\begin{eqnarray}2\pi {R}_{{\rm{cap}}}^{2}(1-\cos \theta )={A}_{0}.\end{eqnarray}$
The bending energy of such a spherical cap reads 9 ) and (10 ) still fulfill all the shape equations and all the BCs except that equation (B5 ) only approximately holds for small fr (see appendix E ). The total free energy of the spherical cap solution 4 ).
$\begin{eqnarray}{F}_{{\rm{sb}}}=2\pi \bar{\kappa }\left(\frac{1}{2}\frac{\bar{\kappa }}{\kappa }+1\right)\left(1-\cos \theta \right)=\left[\frac{\bar{\kappa }}{2\left(2+\bar{\kappa }/\kappa \right)}\right]{c}_{0}^{2}{A}_{0}.\end{eqnarray}$
When a compressive stress fr is applied at the edge of the membrane, the spherical cap solutions expressed by equations ( $\begin{eqnarray}{F}_{\,\rm{s}\,}={F}_{{\rm{sb}}}+{f}_{\,\rm{r}\,}\pi {R}_{{\rm{cap}}}^{2}{\sin }^{2}\theta \end{eqnarray}$
increases linearly with the stress fr due to the boundary energy term equation (3. Results
3.1. Buckling of membranes with different Gaussian curvature moduli
In this section, we study the effects of Gaussian curvature modulus on the buckling of a circular membrane in response to a compressive stress fr. The spontaneous curvature of the membrane is fixed at c0R0=0.1. In addition to the spherical cap solution (figure 2, cyan line), a new branch of solutions, which represent a buckled membrane shape, emerges when the stress fr is beyond a critical value ${f}_{\,\rm{b}\,}^{* }$ (black curve in figure 2). The total free energy of the buckled membrane shape F=Fb+Fl is lower than that of the spherical cap Fs, indicating the occurrence of a buckling transition. Upon further increase of the stress fr to ${f}_{\,\rm{v}\,}^{* }$, the degree of membrane buckling is enhanced until the base radius is narrowed down to almost zero (figure 2(d)), which is termed vesiculation for the rest of the paper. Here it should be emphasized that the buckled membrane shapes deviate from a spherical cap, except at the vesiculation point (see figure 8 and appendix F ). Therefore, our method is different from the vesicle closure simulation [48,49], in which the membrane is assumed to be a spherical cap and closure is driven by a constant edge tension that favors a closed vesicle. When comparing the free energy curves for different Gaussian curvature moduli $\bar{\kappa }$, we find that both the critical stress of membrane buckling ${f}_{\,\rm{b}\,}^{* }$ and the stress of vesiculation ${f}_{\,\rm{v}\,}^{* }$ are reduced for more negative $\bar{\kappa }$ (compare figures 2(a), (b), and (c)). In particular, when $\bar{\kappa }/\kappa =-1.5$, the buckled shape solutions appear even in the absence of compressive stresses and has a lower free energy than the spherical cap solution, indicating a spontaneous buckling of the membrane (figure 2(c)).
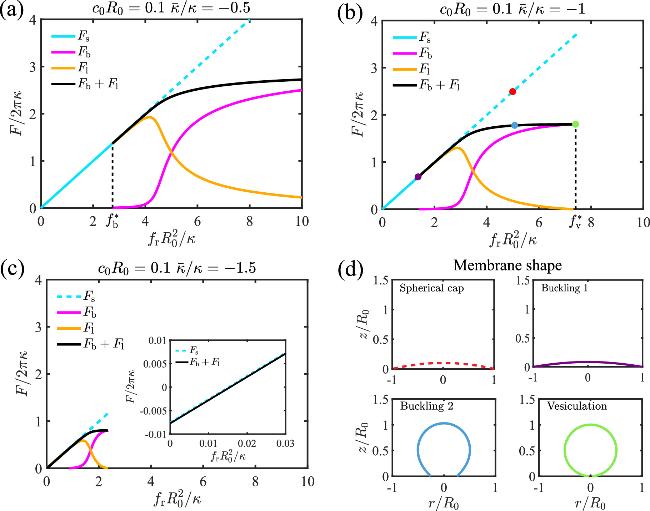
Figure 2. Effect of the stress ${f}_{\,\rm{r}\,}{R}_{0}^{2}/\kappa $ on the free energy F/2πκ of the membrane with a scaled spontaneous curvature c0R0=0.1 based on different Gaussian curvatures (a) $\bar{\kappa }/\kappa =-0.5$, (b) $\bar{\kappa }/\kappa =-1$, and (c) $\bar{\kappa }/\kappa =-1.5$. (d) demonstrates the membrane shapes corresponding to the colored dots in (b). |
To systematically investigate how the Gaussian curvature moduli $\bar{\kappa }$ influence the buckling behavior of a circular membrane, we show the phase diagram of the membrane shape as a function of $\bar{\kappa }$ and the stress fr in figure 3 for different spontaneous curvature c0. When c0R0=0.1 or 0.2, the phase diagram is divided into three parts, separated by the curve of critical buckling stress ${f}_{\,\rm{b}\,}^{* }$ (figures 3(a) and (b), the boundary between the pink region and the grey region), and the curve of critical vesiculation stress ${f}_{\,\rm{v}\,}^{* }$ (figures 3(a) and (b), the boundary between the pink region and the blue region). The area of the grey region, which represents the spherical cap shape, is reduced with increasing spontaneous curvature c0, and vanishes when c0 is large (figure 3(c)). Overall, the results suggest that a negative Gaussian curvature modulus $\bar{\kappa }$ and a positive spontaneous curvature c0 tend to facilitate the buckling by reducing the critical buckling stress ${f}_{\,\rm{b}\,}^{* }$. However, the critical vesiculation stress ${f}_{\,\rm{v}\,}^{* }$ is not strongly influenced by the spontaneous curvature.
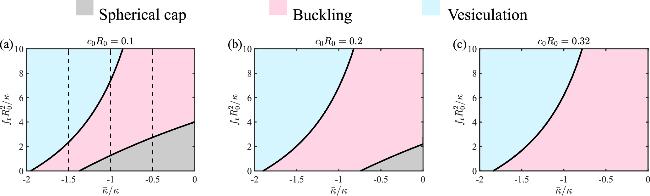
Figure 3. Phase diagram of the membrane shape as a function of the Gaussian curvature modulus $\bar{\kappa }/\kappa $ and the compressive stress ${f}_{\,\rm{r}\,}{R}_{0}^{2}/\kappa $ under different spontaneous curvatures (a) c0R0=0.1, (b) c0R0=0.2 and (c) c0R0=0.32. The dashed lines in (a) represent the cases of figure 2(a), (b) and (c), respectively. |
The effect of the Gaussian curvature modulus on the buckling behavior of membrane can be further reflected on the stress-strain relationship shown in figure 4. We quantify the buckling degree of the membrane by introducing the strain μr=(A0-Ab)/A0, where ${A}_{\rm{b}\,}=\pi {R}_{\,\rm{b}}^{2}$ represents the base area of the buckled membrane. The stress fr(μr=0) defines the critical buckling stress ${f}_{\,\rm{b}\,}^{* }$ and the stress fr(μr=1) defines the critical vesiculation stress ${f}_{\,\rm{v}\,}^{* }$. At the beginning of the buckling when μr is small, the stress fr increases rapidly with μr. At intermediate μr, the stress fr mildly with μr. At the end of the buckling when μr is close to 1, the stress fr rises rapidly again with μr. The vesiculation stress ${f}_{\,\rm{v}\,}^{* }$ can be significantly reduced with more negative $\bar{\kappa }$.
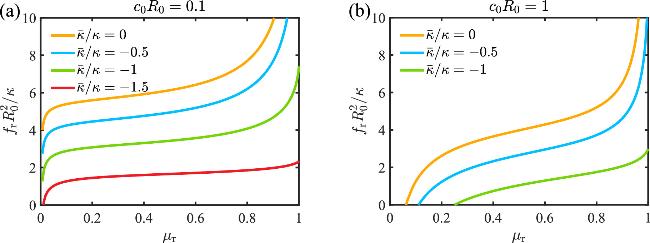
Figure 4. The stress-strain relationship of the buckled membrane shape under different Gaussian curvature moduli, with the spontaneous curvatures (a) c0R0=0.1 and (b) c0R0=1. |
3.2. Buckling of membranes with different spontaneous curvatures
In this section, we investigate the buckling behavior of a circular membrane under different spontaneous curvatures c0R0. The phase diagrams of the membrane shape as a function of the compressive stress fr and the spontaneous curvature c0 are shown in figure 5 for $\bar{\kappa }/\kappa =-0.5$ and $\bar{\kappa }/\kappa =-1$, respectively. It is found that increasing the spontaneous curvature c0 is able to reduce the critical buckling stress ${f}_{\,\rm{b}\,}^{* }$, which is the boundary that separates the grey region from the pink region. When c0 exceeds a critical value, the critical buckling stress ${f}_{\,\rm{b}\,}^{* }$ turns negative (figure 5), indicating spontaneous buckling. In this case, the membrane adopts a buckled shape rather than a spherical cap, even in the absence of compressive stresses. The vesiculation buckling stress ${f}_{\,\rm{v}\,}^{* }$, which is the boundary that separates the blue region from the pink region, is also reduced with increasing c0 (figure 5). The reduction is more dramatic for more negative Gaussian curvature modulus.
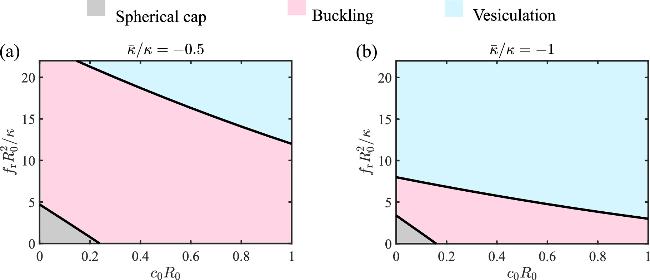
Figure 5. Phase diagram of the membrane constructed by the scaled spontaneous curvature c0R0 and stress ${f}_{\,\rm{r}\,}{R}_{0}^{2}/\kappa $ under different Gaussian curvature moduli (a) $\bar{\kappa }/\kappa =-0.5$ and (b) $\bar{\kappa }/\kappa =-1$. |
3.3. Determination of Gaussian curvature modulus and spontaneous curvature
In this section, we propose a method to determine the Gaussian curvature modulus $\bar{\kappa }$ and the spontaneous curvature c0 simultaneously via a buckling protocol. We assume that the stress-strain relationship of a buckled membrane can be obtained either via molecular dynamics simulation or experiments. In figure 6, we show the stresses at two different strains fr(μr=0.3) and fr(μr=0.6) as a function of the scaled spontaneous curvature c0R0 and the Gaussian curvature modulus $\bar{\kappa }/\kappa $. It is found that the contours of the stress are approximately linear with different slopes for μr=0.3 and μr=0.6. They can be approximately featured by 13 ) and (14 ) to get the approximated value of $\bar{\kappa }/\kappa $ and c0R0. Here we choose two intermediate strains, μr=0.3 and μr=0.6, because (i) these strains are sufficiently large to distinguish the system from the spherical cap solution, confirming that the membrane is in a buckled state, and (ii) the contour profiles can be reasonably approximated by a linear relationship. At very high strains, however, the nonlinearity of the contours becomes evident. The accuracy of the measurement depends on the bandwidth between different contours. In principle, the Gaussian curvature modulus $\bar{\kappa }$ and the spontaneous curvature c0 can be determined with a very high accuracy when multiple points on the stress-strain relationship are used.
$\begin{eqnarray}\frac{\bar{\kappa }}{\kappa }=1.12{c}_{0}{R}_{0}+{g}_{0}(\frac{{f}_{\,\rm{r}\,0}{R}_{0}^{2}}{\kappa }),\end{eqnarray}$
$\begin{eqnarray}\frac{\bar{\kappa }}{\kappa }=0.8{c}_{0}{R}_{0}+{g}_{1}(\frac{{f}_{\,\rm{r}\,1}{R}_{0}^{2}}{\kappa }),\end{eqnarray}$
where fr0=fr(μr=0.3) and fr1=fr(μr=0.6) are the stresses at two particular strains. The intercept functions g0(x) and g1(x) can be well fitted by a quadratic function (figures 6(b) and 6(d)). Therefore, by measuring the buckled stress fr0 and fr1 at μr=0.3 and μr=0.6, we can solve equations (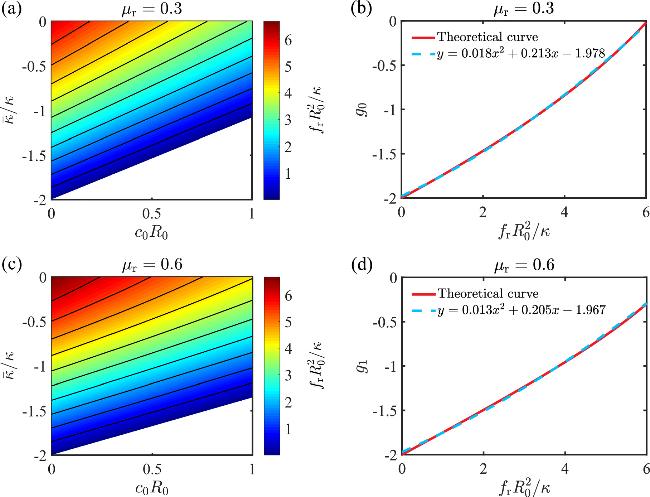
Figure 6. Measuring the spontaneous curvature c0R0 and the Gaussian curvature modulus $\bar{\kappa }/\kappa $ of a membrane using the stress-strain relationship in a buckling process. (a, b) The buckling stress ${f}_{\,\rm{r}\,}{R}_{0}^{2}/\kappa $ at a buckling strain μr=0.3 as a function of c0R0 and $\bar{\kappa }/\kappa $ in (a). The contour lines approximately have the same slope and can be fitted with a linear function $\bar{\kappa }/\kappa =1.12{c}_{0}{R}_{0}+{g}_{0}({f}_{\,\rm{r}\,0}{R}_{0}^{2}/\kappa )$, where the intercepts g0(x) can be approximately featured by a quadratic function shown in (b). (c, d) The same as in (a, b) except that the strain μr=0.6. The contour lines can be characterized by a linear function $\bar{\kappa }/\kappa =0.80{c}_{0}{R}_{0}+{g}_{1}({f}_{\,\rm{r}\,1}{R}_{0}^{2}/\kappa )$, where the intercepts g1(x) can be fitted by a quadratic function shown in (d). The empty areas in (a) and (c) indicate regions where a negative stress is required to maintain a buckled shape at strains μr=0.3 and μr=0.6. In these regions, spontaneous buckling would occur, preventing a well-defined transition from a spherical cap to a buckled shape. Since our focus is on using buckling as a protocol to measure $\bar{\kappa }/\kappa $ and c0R0, these cases are beyond the scope of our study. |
4. Discussion
Membrane properties such as bending rigidity, spontaneous curvature, and Gaussian bending rigidity can be measured using various experimental and computational techniques. Bending rigidity κ is commonly determined through tether pulling experiment, where a thin membrane tube (tether) is pulled from a vesicle or cell membrane using an external force, or through fluctuation spectroscopy, which examines thermal undulations of membranes to extract elastic parameters [38]. Spontaneous curvature c0, which arises from lipid asymmetry or interactions, can be inferred from the axial force to maintain the membrane as a tubular shape [38]. The Gaussian curvature modulus $\bar{\kappa }$, which governs topological transitions, is more challenging to measure directly but can be inferred from vesicle fusion and fission dynamics, membrane budding experiments, or computational simulations that model topological changes such as vesicle closure [48,49]. These techniques, whether experimental or computational, offer complementary approaches to quantify membrane mechanical properties. In this work, we study the buckling of a circular membrane under compressive stresses. The buckled membrane shape is obtained by numerically solving the variational equations associated with the membrane's bending energy. Based on the stress-strain relationship, we propose a method to measure the Gaussian curvature modulus by determining the buckling stress at two specific strain values. Our method differs from the vesicle closure simulation [48,49] in three key aspects: (i) while they assumed a constant edge tension, we varied the edge stress to induce membrane buckling; (ii) they constrained the membrane shape within a spherical cap, whereas we determined the shape by solving the variational equations, which is revealed to deviate from a spherical cap; and (iii) they took into account the thermal fluctuations of the membrane shape, which are particularly significant in molecular dynamics simulations of membranes with a relatively small number of lipid molecules [38], whereas we did not. When applying our method to simulate the buckling process or conducting experiments, special attention must be given to fluctuations, as they introduce uncertainties in the measurement of strains and stresses. In computer simulations, a circular edge can be maintained by freezing the positions of lipid molecules at the boundary. In experiments, however, the diffusive motion of lipid molecules at the edge may cause deviations from a perfect circular shape. To mitigate this issue, a possible approach is to construct a fence-like structure with a fixed circular shape. The membrane is placed inside a fence in a relatively compact manner to limit edge fluctuations, while the fence's radius remains adjustable to allow the controlled compression of the edge. Understanding how these fluctuations influence the stress-strain relationship will be the focus of our future work.
In our spontaneous curvature model of membranes, the bending energy is expressed in the form of equation (3 ), where the elastic properties of the membrane are fully characterized by three phenomenological parameters: the mean curvature modulus κ, the Gaussian curvature modulus $\bar{\kappa }$, and the spontaneous curvature c0. Such a characterization is based on the assumption that there are no lipids exchange between the two leaflets, therefore the composition of the lipids and the number of lipids for the two leaflets are fixed. Furthermore, in the presence of multi-component membrane, it is required that different components are well mixed in a homogeneous way.
In reality, membrane behavior can be more complex than predicted by the spontaneous model employed in our work [66]. For instance, as the system's temperature gradually increases, a spherical vesicle tends to undergo a sequence of shape transformations, ultimately leading to vesiculation, where two spheres remain connected by a narrow neck. In this case, the spontaneous curvature model fails to capture the discontinuous transition from a pear-shaped vesicle to vesiculation [40]. To accurately describe this sequence of shape transformations, an area-difference elasticity (ADE) model has been developed. This model incorporates an additional free-energy term,
$\begin{eqnarray}{f}_{{\rm{n}}{\rm{l}}}=\frac{{\kappa }_{{\rm{n}}{\rm{l}}}}{2}{(\bar{c}-{\bar{c}}_{0})}^{2},\end{eqnarray}$
which arises from differential stresses between the two leaflets of the bilayer [40,67]. Here, the surface-averaged mean curvature, $\begin{eqnarray*}\bar{c}=\frac{1}{A}\int ({c}_{1}+{c}_{2})\,\,\rm{d}\,A,\end{eqnarray*}$
is related to the area difference between the two leaflets. The differential stress curvature ${\bar{c}}_{0}$ and the nonlocal bending rigidity κnl are two additional parameters characterizing the membrane's mechanical properties. When lipid density asymmetry exists between the two leaflets, the contribution made by this term can not be neglected [67]. The reason lies in that this term introduces a nonlocal effect, transforming the variational equations of the ADE model into a set of integro-differential equations. Investigating its impact on the buckling process of a circular membrane will be our future work.5. Conclusion
In this paper, we aim to study the role of Gaussian curvature modulus $\bar{\kappa }$ and spontaneous curvature c0 in membrane buckling under the free-hinge boundary condition by investigating the buckling behaviors of a circular membrane under radially oriented compressive stress. When c0 and $| \bar{\kappa }| $ are small, in the absence of the stress, the circular membrane typically adopts a spherical cap shape, which remains unchanged until the stress exceeds a critical value. Then the membrane would buckle in response to the stress and the base radius is reduced to almost zero with increasing stress. A more negative Gaussian curvature modulus $\bar{\kappa }$ is able to reduce the critical buckling stress to negative values, such that buckling would occur spontaneously in the absence of compressive stresses.
We have proposed a method to determine the Gaussian curvature modulus and the spontaneous curvature simultaneously for molecular dynamics simulations and experiments. The method uses the stress-strain relationship of a circular membrane and compare the measured stress with the calculated stress at different strains. The challenge of this method for molecular dynamics simulations is to impose the free-hinge boundary condition which can not be fulfilled by the typical periodic boundary condition in the simulation box. Implementing this method in molecular dynamics simulations will be our future work.
Acknowledgments
We acknowledge the financial support from the National Natural Science Foundation of China under Grant Nos. 12174323 and 12474199, Fundamental Research Funds for Central Universities of China under Grant No. 20720240144 (RM), and 111 project B16029.
Appendix A. Derivation of the membrane shape equations
In this section, we provide the mathematical details of how to derive the equations we used to solve the buckled membrane shape. The effective free energy $\bar{F}$ in equation (7 ) is a functional of the membrane shape. The integrand ${ \mathcal L }$ inside the integral can be written as 2 ). By applying the variational method to the functional $\bar{F}$, we obtain A2 ) vanish, we obtain the following shape equations 2 ). Note that by letting $\partial { \mathcal L }/\partial h=0$, instead of obtaining a shape equation, we derive a conserved quantity
$\begin{eqnarray}\begin{array}{rcl}{ \mathcal L } & =& \frac{1}{2}{\left(\frac{{\psi }^{{\prime} }}{h}+\frac{\sin \psi }{r}-{c}_{0}\right)}^{2}rh+\bar{\kappa }{\psi }^{{\prime} }\sin \psi /\kappa +\bar{\sigma }rh\\ & & +{\bar{f}}_{\,\rm{r}\,}rh\cos \psi +\gamma \left({r}^{{\prime} }-h\cos \psi \right)+\eta \left({z}^{{\prime} }+h\sin \psi \right),\end{array}\end{eqnarray}$
where $\bar{\sigma }=\sigma /\kappa $, $\bar{{f}_{\rm{r}\,}}={f}_{\,\rm{r}}/\kappa $, and γ(u) and η(u) are Lagrangian multipliers to enforce the geometric relations in equation ( $\begin{eqnarray}\begin{array}{rcl}\frac{\delta F}{2\pi \kappa } & =& {\displaystyle \int }_{0}^{1}\,\rm{d}\,u\left[\left(\frac{\partial { \mathcal L }}{\partial \psi }-\frac{\rm{d}\,}{\,\rm{d}u}\frac{\partial { \mathcal L }}{\partial {\psi }^{{\prime} }}\right)\delta \psi +\left(\frac{\partial { \mathcal L }}{\partial r}-\frac{\rm{d}\,}{\,\rm{d}u}\frac{\partial { \mathcal L }}{\partial {r}^{{\prime} }}\right)\delta r\right.\\ & & \left.+\left(\frac{\partial { \mathcal L }}{\partial z}-\frac{\rm{d}\,}{\,\rm{d}u}\frac{\partial { \mathcal L }}{\partial {z}^{{\prime} }}\right)\delta z+\frac{\partial { \mathcal L }}{\partial \gamma }\delta \gamma +\frac{\partial { \mathcal L }}{\partial \eta }\delta \eta +\frac{\partial { \mathcal L }}{\partial h}\delta h\right]\\ & & {\left.+\frac{\partial { \mathcal L }}{\partial {\psi }^{{\prime} }}\delta \psi \right|}_{u=0}^{u=1}+{\left.\frac{\partial { \mathcal L }}{\partial {r}^{{\prime} }}\delta r\right|}_{u=0}^{u=1}+{\left.\frac{\partial { \mathcal L }}{\partial {z}^{{\prime} }}\delta z\right|}_{u=0}^{u=1}.\end{array}\end{eqnarray}$
If the bulk terms of equation ( $\begin{eqnarray}\begin{array}{rcl}{\psi }^{{\prime\prime} } & =& \frac{{h}^{2}\sin \psi \cos \psi }{{r}^{2}}-\frac{{\psi }^{{\prime} }}{r}h\cos \psi -{\bar{f}}_{\,\rm{r}\,}{h}^{2}\sin \psi \\ & & +\frac{\gamma }{r}{h}^{2}\sin \psi +\frac{\eta }{r}{h}^{2}\cos \psi ,\end{array}\end{eqnarray}$
$\begin{eqnarray}{\gamma }^{{\prime} }=h\left\{\frac{1}{2}\left[{\left(\frac{{\psi }^{{\prime} }}{h}-{c}_{0}\right)}^{2}-{\left(\frac{\sin \psi }{r}\right)}^{2}\right]+\bar{\sigma }+{\bar{f}}_{\,\rm{r}\,}\cos \psi \right\},\end{eqnarray}$
$\begin{eqnarray}{\eta }^{{\prime} }=0,\end{eqnarray}$
as well as the two geometric relations in equation ( $\begin{eqnarray}\begin{array}{rcl}H & =& \frac{r}{2}\left[{\left(\frac{{\psi }^{{\prime} }}{h}\right)}^{2}-{\left(\frac{\sin \psi }{r}-{c}_{0}\right)}^{2}\right]-\bar{\sigma }r-\bar{{f}_{\,\rm{r}\,}}r\cos \psi \\ & & +\gamma \cos \psi -\eta \sin \psi =0,\end{array}\end{eqnarray}$
which will be used as a boundary condition.Appendix B. Derivation of the boundary conditions
The boundary conditions can be derived by setting the boundary terms in equation (A2 ) to be zero. In particular, at the membrane tip u=0, we fix the angle and the radius by requiring B1 ) and (B2 ), implying that
$\begin{eqnarray}\psi (u=0)=0,\quad \quad r(u=0)=0.\end{eqnarray}$
As for the boundary term about Δz, we let the conjugate term $\partial { \mathcal L }/\partial {z}^{{\prime} }=0$, which means $\begin{eqnarray}\eta (u=0)=0.\end{eqnarray}$
Meanwhile we also require H(0)=0, together with ( $\begin{eqnarray}\gamma (u=0)=0.\end{eqnarray}$
At the membrane base u=1, we impose $\partial { \mathcal L }/\partial {\psi }^{{\prime} }=0$, which gives the free-hinge boundary condition equation (8 ). It can be explicitly written as
$\begin{eqnarray}{\left[\kappa \left(\frac{{\psi }^{{\prime} }}{h}+\frac{\sin \psi }{r}-{c}_{0}\right)+\bar{\kappa }\left(\frac{\sin \psi }{r}\right)\right]}_{u=1}=0.\end{eqnarray}$
We also impose $\delta L/\delta {r}^{{\prime} }=0$, which is equivalent to $\begin{eqnarray}\gamma (u=1)=0.\end{eqnarray}$
This means that the base radius r(u=1) is determined by the equations provided the compressive stress fr is given. As for the boundary term about Δz, we impose Δz (u=1)=0, which is to fix the z-coordinate $\begin{eqnarray}z(u=1)=0.\end{eqnarray}$
Finally, we impose the incompressibility condition by requiring that the membrane area is a constant 6 ), and a function a(u) that satisfies the equation
$\begin{eqnarray}2\pi {\int }_{0}^{1}rh\,\rm{d}\,u=\pi {R}_{0}^{2}.\end{eqnarray}$
This is achieved by introducing the Lagrangian multiplier σ in equation ( $\begin{eqnarray}{a}^{{\prime} }=2\pi rh,\end{eqnarray}$
with two boundary conditions $\begin{eqnarray}a(u=0)=0,\quad \quad a(u=1)=\pi {R}_{0}^{2}.\end{eqnarray}$
In summary, equations (2 ), (A3 ), (A4 ), (A5 ) and (B8 ) make up the full set of shape equations for the buckling problem. They are equivalent to 7 first order ordinary differential equations. In addition, there are two unknown parameters h and σ, which represent the total arclength and the membrane tension, respectively. Together with the nine boundary conditions given by equations (B1 )-(B6 ), and (B9 ), we construct a well-defined boundary value problem.
Appendix C. Interpretation of the boundary conditions
The introduction of the Lagrange multipliers significantly simplifies the shape equations. However, their physical implications are still obscure. To understand the physical meaning of the boundary conditions associated with the Lagrange multipliers, we solve η and γ from equations (A3 ) and (A6 ), and rearrange them into the following forms: B2 ) and the shape equation (A5 ) together imply that η(u)=0 for the entire interval u ∈ [0, 1], leading to η(u=1)=0. This condition, together with Eq.(B5 ), ensures that the right-hand sides of equations (C1 ) and (C2 ) vanish, enforcing the force balance conditions in both the normal and the tangential directions at the edge. This is consistent with the previous studies on general shape equations for membranes with an open edge [39,64].
$\begin{eqnarray}\frac{\eta \cos \psi +\gamma \sin \psi }{r}=\frac{1}{h}\frac{\rm{d}\,}{\,\rm{d}u}\left(\frac{{\psi }^{{\prime} }}{h}+\frac{\sin \psi }{r}\right)+\bar{{f}_{\,\rm{r}\,}}\sin \psi ,\end{eqnarray}$
and $\begin{eqnarray}\begin{array}{rcl}\frac{\eta \sin \psi -\gamma \cos \psi }{r} & =& \frac{1}{2}\left[-{\left(\frac{{\psi }^{{\prime} }}{h}\right)}^{2}+{\left(\frac{\sin \psi }{r}-{c}_{0}\right)}^{2}\right]\\ & & +\bar{\sigma }+{\bar{f}}_{\,\rm{r}\,}\cos \psi .\end{array}\end{eqnarray}$
The boundary condition (Here it should be noted that in the presence of edge tension γ, the boundary stress ${\bar{f}}_{\,\rm{r}\,}$ in equations (C1 ) and (C2 ) should be modified as fr+γ/r. This equation implies that the edge tension plays an important role in the late stage of buckling when the base radius Rb is small. Therefore, the critical vesiculation stress ${f}_{\,\rm{v}\,}^{* }$ will be reduced. Nevertheless, the influence on the critical buckling stress ${f}_{\,\rm{b}\,}^{* }$ by the edge tension is still trivial if γ/R0 is small.
Appendix D. Numerical method to calculate the stress-strain relationship
In order to obtain the stress-strain relationship and the corresponding buckled membrane shapes, the boundary value problem can be numerically solved with a given stress fr. We use the boundary value problem solver bvp5c in MATLAB to solve the problem. However, the solver requires a guess of the solution which makes it difficult to obtain the correct solution for an arbitrary value of fr if the guessed solution is far from the correct solution. To overcome this issue, we make the stress fr an unknown parameter and introduce another boundary condition
$\begin{eqnarray}r(u=1)={R}_{\,\rm{b}\,},\end{eqnarray}$
to fix the base radius. We always begin with a nearly flat membrane, defined by r(u)=ε+Rbu and z(u)=z0(1-u2), where ε=0.0001 is chosen so as to prevent singularities at the membrane tip, and z0=0.01 slightly elevates the membrane. Then we iteratively solve the problem by reducing the base radius Rb with a small increment ΔRb until Rb is almost zero. The solution for Rb+ΔRb is used for the guessed solution for Rb. In this way, we obtain the stress fr for different strain ${\mu }_{\rm{r}\,}=1-{R}_{\,\rm{b}}^{2}/{R}_{0}^{2}$, i.e., the stress-strain relationship.Appendix E. Proof of the spherical-cap solution
For a spherical cap with a radius of Rcap and a spanning angle θ, the solutions read 9 ) and (10 ). The two Lagrangian multipliers can be obtained by substituting equations (E1 )-(E3 ) into equations (A3 ) and (A6 ), which gives E5 ), then η(u)=0 is satisfied. In the absence of boundary stress (i.e., ${\bar{f}}_{\,\rm{r}\,}=0$), equation (E4 ) implies γ=0, meaning the spherical cap solutions always satisfy all the shape equations and boundary conditions. However, when a boundary stress is applied, the spherical cap solutions (E1 )-(E6 ) satisfy all the shape equations and boundary conditions, except that $\gamma (u)={R}_{\,\rm{cap}\,}{\bar{f}}_{\,\rm{r}\,}\sin (u\theta )$ only approximately satisfies the boundary condition γ(u=1)=0 in equation (B5 ). This approximation holds for small ${\bar{f}}_{\,\rm{r}\,}$ and θ, which is generally valid when fr remains below the critical stress ${f}_{\,\rm{b}\,}^{* }$. To demonstrate that, we compare the spherical cap solution with the exact numerical solutions and find a good agreement between them in figure 7.
$\begin{eqnarray}r(u)={R}_{\,\rm{cap}\,}\sin (u\theta ),\end{eqnarray}$
$\begin{eqnarray}z(u)={R}_{\,\rm{cap}\,}[\cos (u\theta )-\cos (\theta )],\end{eqnarray}$
$\begin{eqnarray}\psi (u)=u\theta ,h(u)={R}_{\,\rm{cap}\,}\theta .\end{eqnarray}$
where Rcap and θ are given by equations ( $\begin{eqnarray}\gamma (u)=\frac{1}{2}\left[2{R}_{\,\rm{cap}\,}{\bar{f}}_{\,\rm{r}\,}-\left(2{c}_{0}-{c}_{0}^{2}{R}_{\rm{cap}\,}-2{R}_{\,\rm{cap}}\bar{\sigma }\right)\right]\sin (u\theta ),\end{eqnarray}$
$\begin{eqnarray}\eta (u)=\frac{1}{2}\left(2{c}_{0}-{c}_{0}^{2}{R}_{\rm{cap}\,}-2{R}_{\,\rm{cap}}\bar{\sigma }\right){\sin }^{2}(u\theta ).\end{eqnarray}$
If we let the reduced membrane tension $\begin{eqnarray}\bar{\sigma }=\frac{2{c}_{0}-{c}_{0}^{2}{R}_{\rm{cap}\,}}{2{R}_{\,\rm{cap}}},\end{eqnarray}$
in equation (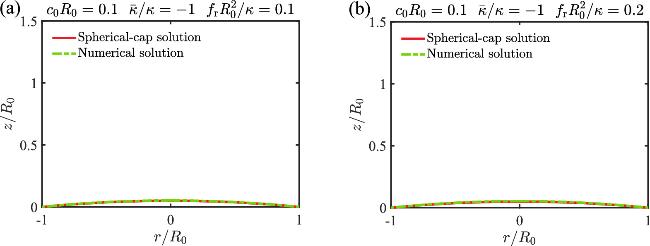
Figure 7. Comparison between the spherical cap solution and the exact numerical solution for boundary stresses below the critical buckling stress (${f}_{\rm{r}\,}\lt {f}_{\,\rm{b}}^{* }$). (a) and (b) show results for different values of fr. |
Appendix F. Analysis of the buckled shape
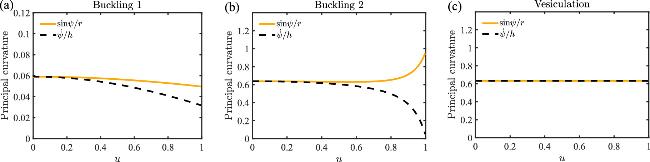
In figure 2(d), we show the buckled membrane shapes for c0R0=0.1 and $\bar{\kappa }/\kappa =-1$, and stress that the buckled shapes deviate from a spherical cap except the one at vesiculation point. To tell that, we show the two principal curvatures of the shapes in figure 8, where deviation between the two principal curvatures are observed for the first two buckled shapes. At the vesiculation point, the buckled shape becomes a sphere such that the two principal curvatures overlap and remain constant.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Figure 8. Principal curvatures of buckled membrane shapes. (a-c) correspond to the three buckled solutions shown in figure 2(d). |